

L-2-Hydroxyglutarate: an epigenetic modifier and putative oncometabolite in renal cancer
- PMID: 25182153
- PMCID: PMC4286872
- DOI: 10.1158/2159-8290.CD-13-0696
L-2-Hydroxyglutarate: an epigenetic modifier and putative oncometabolite in renal cancer
Abstract
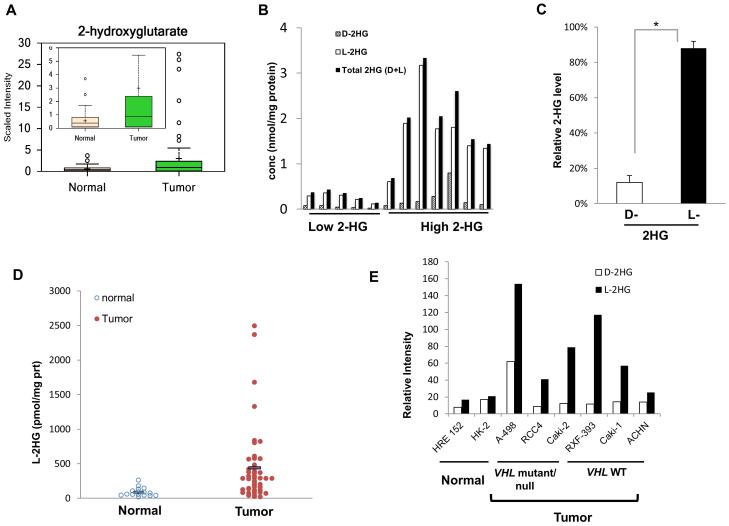
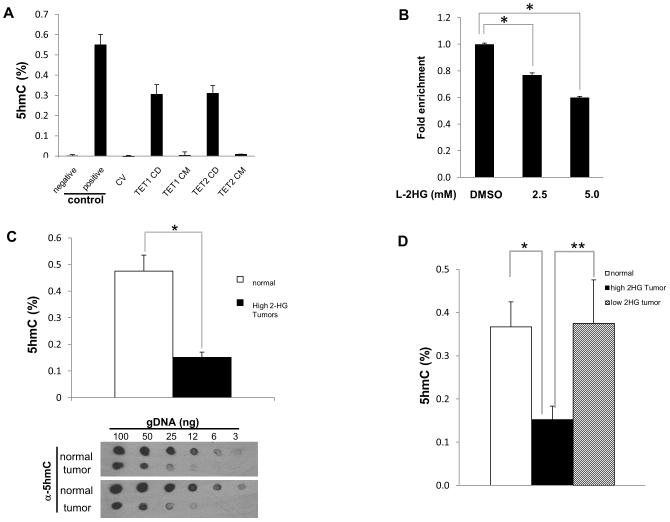
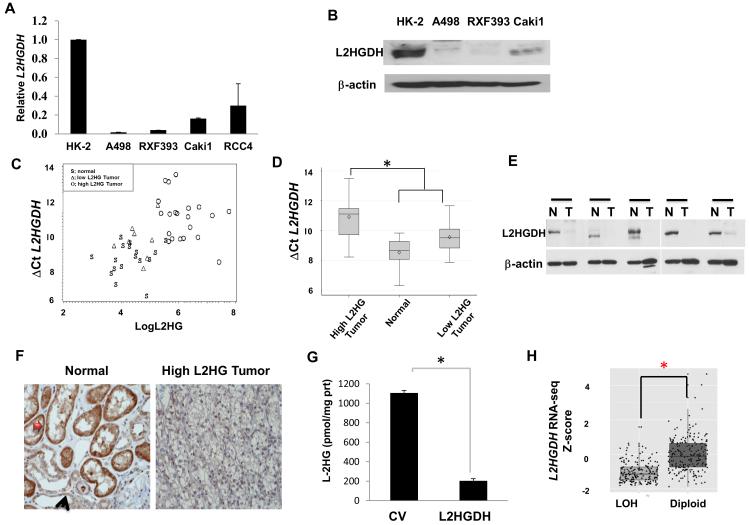
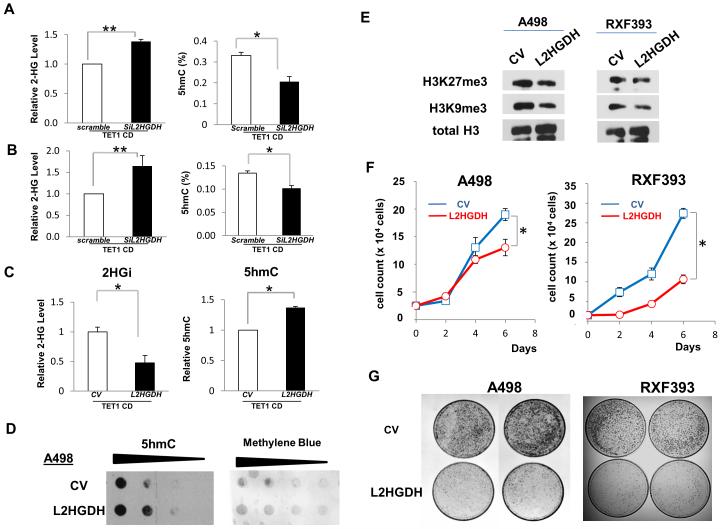
Through unbiased metabolomics, we identified elevations of the metabolite 2-hydroxyglutarate (2HG) in renal cell carcinoma (RCC). 2HG can inhibit 2-oxoglutaratre (2-OG)-dependent dioxygenases that mediate epigenetic events, including DNA and histone demethylation. 2HG accumulation, specifically the d enantiomer, can result from gain-of-function mutations of isocitrate dehydrogenase (IDH1, IDH2) found in several different tumors. In contrast, kidney tumors demonstrate elevations of the l enantiomer of 2HG (l-2HG). High-2HG tumors demonstrate reduced DNA levels of 5-hydroxymethylcytosine (5hmC), consistent with 2HG-mediated inhibition of ten-eleven translocation (TET) enzymes, which convert 5-methylcytosine (5mC) to 5hmC. l-2HG elevation is mediated in part by reduced expression of l-2HG dehydrogenase (L2HGDH). L2HGDH reconstitution in RCC cells lowers l-2HG and promotes 5hmC accumulation. In addition, L2HGDH expression in RCC cells reduces histone methylation and suppresses in vitro tumor phenotypes. Our report identifies l-2HG as an epigenetic modifier and putative oncometabolite in kidney cancer.
Significance: Here, we report elevations of the putative oncometabolite l-2HG in the most common subtype of kidney cancer and describe a novel mechanism for the regulation of DNA 5hmC levels. Our findings provide new insight into the metabolic basis for the epigenetic landscape of renal cancer.
©2014 American Association for Cancer Research.
Figures
References
-
- Pollard PJ, Briere JJ, Alam NA, Barwell J, Barclay E, Wortham NC, et al. Accumulation of Krebs cycle intermediates and over-expression of HIF1alpha in tumours which result from germline FH and SDH mutations. Human molecular genetics. 2005;14:2231–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
