

Balanced changes in Ca buffering by SERCA and troponin contribute to Ca handling during β-adrenergic stimulation in cardiac myocytes
- PMID: 25183792
- PMCID: PMC4240166
- DOI: 10.1093/cvr/cvu201
Balanced changes in Ca buffering by SERCA and troponin contribute to Ca handling during β-adrenergic stimulation in cardiac myocytes
Abstract
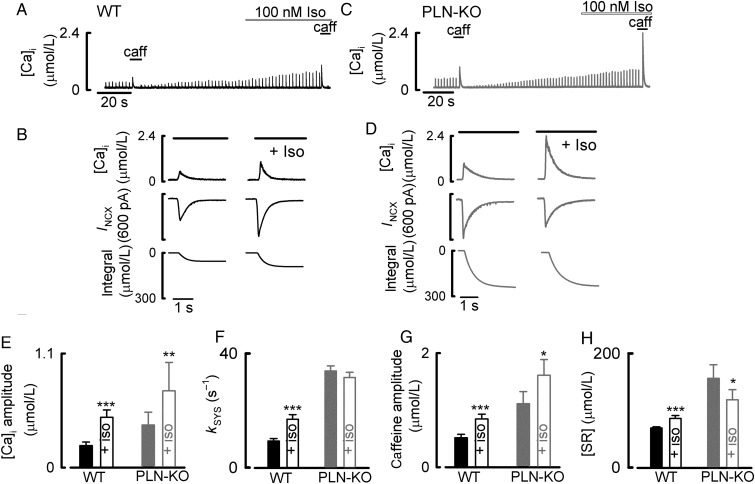
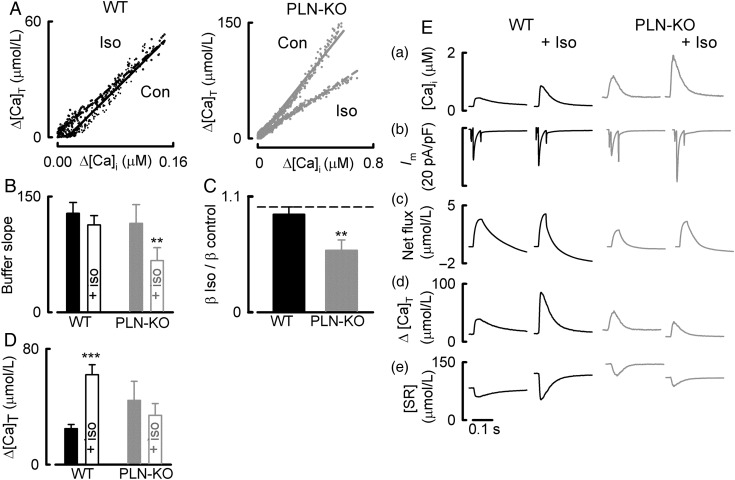
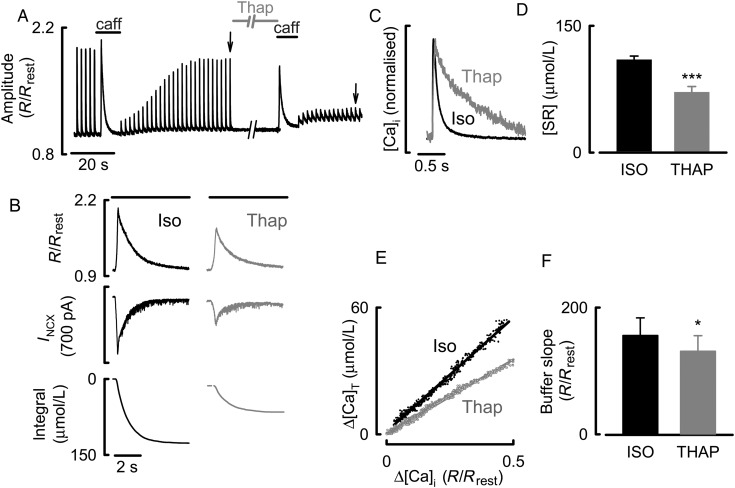
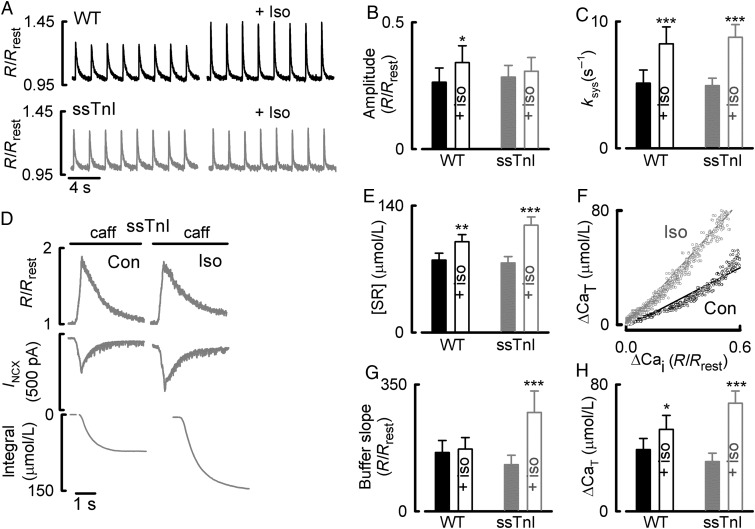
Aims: During activation of cardiac myocytes, less than 1% of cytosolic Ca is free; the rest is bound to buffers, largely SERCA, and troponin C. Signalling by phosphorylation, as occurs during β-adrenergic stimulation, changes the Ca-binding affinity of these proteins and may affect the systolic Ca transient. Our aim was to determine the effects of β-adrenergic stimulation on Ca buffering and to differentiate between the roles of SERCA and troponin.
Methods and results: Ca buffering was studied in cardiac myocytes from mice: wild-type (WT), phospholamban-knockout (PLN-KO), and mice expressing slow skeletal troponin I (ssTnI) that is not protein kinase A phosphorylatable. WT cells showed no change in Ca buffering in response to the β-adrenoceptor agonist isoproterenol (ISO). However, ISO decreased Ca buffering in PLN-KO myocytes, presumably unmasking the role of troponin. This effect was confirmed in WT cells in which SERCA activity was blocked with the application of thapsigargin. In contrast, ISO increased Ca buffering in ssTnI cells, presumably revealing the effect of an increase in Ca binding to SERCA.
Conclusions: These data indicate the individual roles played by SERCA and troponin in Ca buffering during β-adrenergic stimulation and that these two buffers effectively counterbalance each other so that Ca buffering remains constant during β-adrenergic stimulation, a factor which may be physiologically important. This study also emphasizes the importance of taking into account Ca buffering, particularly in disease states where Ca binding to myofilaments or SERCA may be altered.
Keywords: Buffering; Calcium; Phospholamban; SERCA; Troponin.
© The Author 2014. Published by Oxford University Press on behalf of the European Society of Cardiology.
Figures
Similar articles
-
Dissociation of FKBP12.6 from ryanodine receptor type 2 is regulated by cyclic ADP-ribose but not beta-adrenergic stimulation in mouse cardiomyocytes.Cardiovasc Res. 2009 Nov 1;84(2):253-62. doi: 10.1093/cvr/cvp212. Epub 2009 Jul 3. Cardiovasc Res. 2009. PMID: 19578067 Free PMC article.
-
S-Nitrosylation of Calcium-Handling Proteins in Cardiac Adrenergic Signaling and Hypertrophy.Circ Res. 2015 Oct 9;117(9):793-803. doi: 10.1161/CIRCRESAHA.115.307157. Epub 2015 Aug 10. Circ Res. 2015. PMID: 26259881 Free PMC article.
-
Phosphorylation of phospholamban and troponin I in beta-adrenergic-induced acceleration of cardiac relaxation.Am J Physiol Heart Circ Physiol. 2000 Mar;278(3):H769-79. doi: 10.1152/ajpheart.2000.278.3.H769. Am J Physiol Heart Circ Physiol. 2000. PMID: 10710345
-
Regulation of phospholamban and troponin-I phosphorylation in the intact rat cardiomyocytes by adrenergic and cholinergic stimuli: roles of cyclic nucleotides, calcium, protein kinases and phosphatases and depolarization.Mol Cell Biochem. 1995 Aug-Sep;149-150:103-26. doi: 10.1007/BF01076569. Mol Cell Biochem. 1995. PMID: 8569720 Review.
-
Calcium Buffering in the Heart in Health and Disease.Circulation. 2019 May 14;139(20):2358-2371. doi: 10.1161/CIRCULATIONAHA.118.039329. Circulation. 2019. PMID: 31082292 Free PMC article. Review.
Cited by
-
The Control of Diastolic Calcium in the Heart: Basic Mechanisms and Functional Implications.Circ Res. 2020 Jan 31;126(3):395-412. doi: 10.1161/CIRCRESAHA.119.315891. Epub 2020 Jan 30. Circ Res. 2020. PMID: 31999537 Free PMC article. Review.
-
High tension in sarcomeres hinders myocardial relaxation: A computational study.PLoS One. 2018 Oct 4;13(10):e0204642. doi: 10.1371/journal.pone.0204642. eCollection 2018. PLoS One. 2018. PMID: 30286135 Free PMC article.
-
Cardiac inotropy, lusitropy, and Ca2+ handling with major metabolic substrates in rat heart.Pflugers Arch. 2016 Nov;468(11-12):1995-2006. doi: 10.1007/s00424-016-1892-8. Epub 2016 Oct 28. Pflugers Arch. 2016. PMID: 27796576 Free PMC article.
-
Estimating the probability of early afterdepolarizations and predicting arrhythmic risk associated with long QT syndrome type 1 mutations.Biophys J. 2023 Oct 17;122(20):4042-4056. doi: 10.1016/j.bpj.2023.09.001. Epub 2023 Sep 12. Biophys J. 2023. PMID: 37705243 Free PMC article.
-
Neuronal nitric oxide synthase modulation of intracellular Ca2+ handling overrides fatty acid potentiation of cardiac inotropy in hypertensive rats.Pflugers Arch. 2017 Oct;469(10):1359-1371. doi: 10.1007/s00424-017-1991-1. Epub 2017 May 22. Pflugers Arch. 2017. PMID: 28534086
References
-
- Bers DM. Excitation-Contraction Coupling and Cardiac Contractile Force. Dordrecht/Boston/London: Kluwer Academic Publishers; 2001.
-
- Eisner DA, Choi HS, Díaz ME, O'Neill SC, Trafford AW. Integrative analysis of calcium cycling in cardiac muscle. Circ Res. 2000;87:1087–1094. - PubMed
-
- Trafford AW, Díaz ME, Eisner DA. A novel, rapid and reversible method to measure Ca buffering and timecourse of total sarcoplasmic reticulum Ca content in cardiac ventricular myocytes. Pflügers Archiv. 1999;437:501–503. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- CH/2000/04/BHF_/British Heart Foundation/United Kingdom
- FS/10/71/28563/BHF_/British Heart Foundation/United Kingdom
- R01 HL 022231/HL/NHLBI NIH HHS/United States
- PG/10/89/28630/BHF_/British Heart Foundation/United Kingdom
- FS/09/002/26487/BHF_/British Heart Foundation/United Kingdom
- PG/12/89/29970/BHF_/British Heart Foundation/United Kingdom
- P01 HL 062426/HL/NHLBI NIH HHS/United States
- P01 HL062426/HL/NHLBI NIH HHS/United States
- FS/12/57/29717/BHF_/British Heart Foundation/United Kingdom
- FS/09/036/27823/BHF_/British Heart Foundation/United Kingdom
- FS/09/002/BHF_/British Heart Foundation/United Kingdom
- FS/12/57/BHF_/British Heart Foundation/United Kingdom
- PG/11/16/28777/BHF_/British Heart Foundation/United Kingdom
- PG/09/062/27872/BHF_/British Heart Foundation/United Kingdom
- PG/11/16/BHF_/British Heart Foundation/United Kingdom
- PG/08/078/25593/BHF_/British Heart Foundation/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
