

Diverse bacterial microcompartment organelles
- PMID: 25184561
- PMCID: PMC4187681
- DOI: 10.1128/MMBR.00009-14
Diverse bacterial microcompartment organelles
Abstract
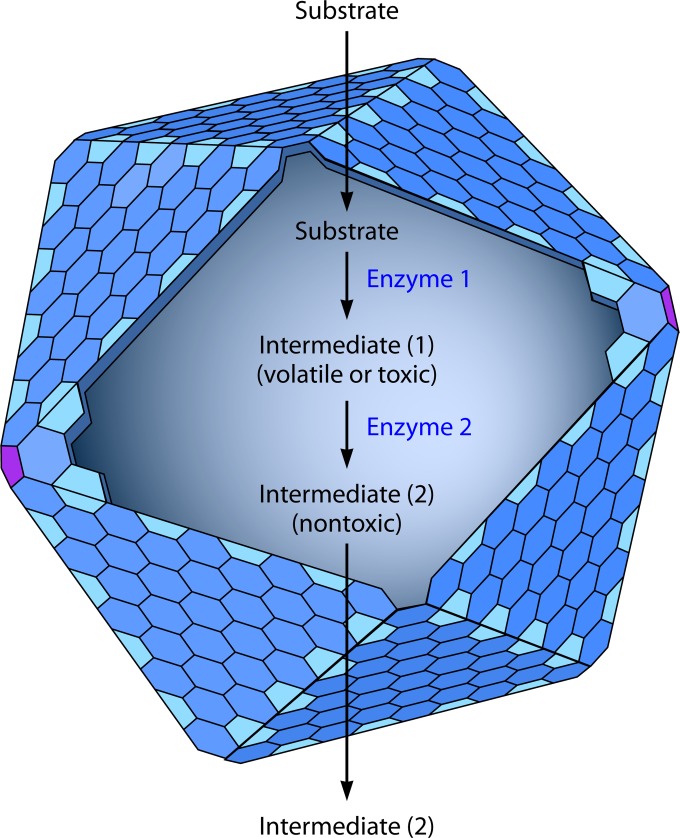
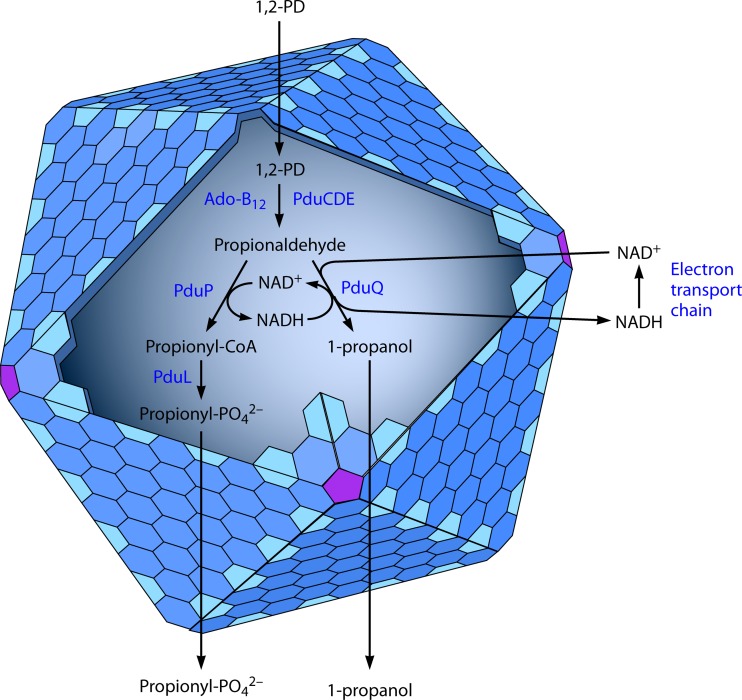
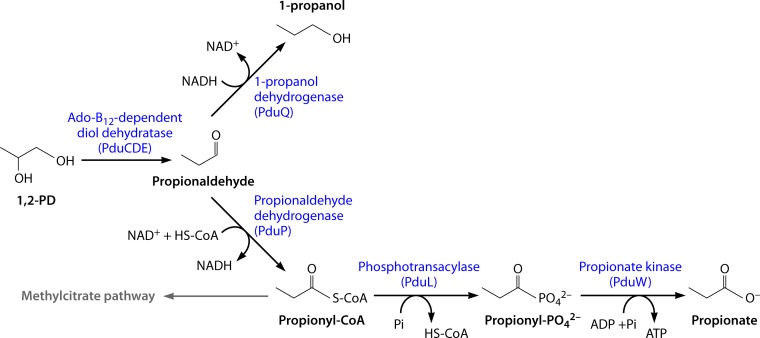
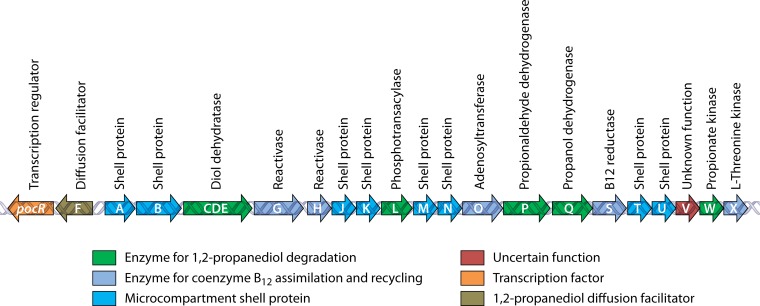
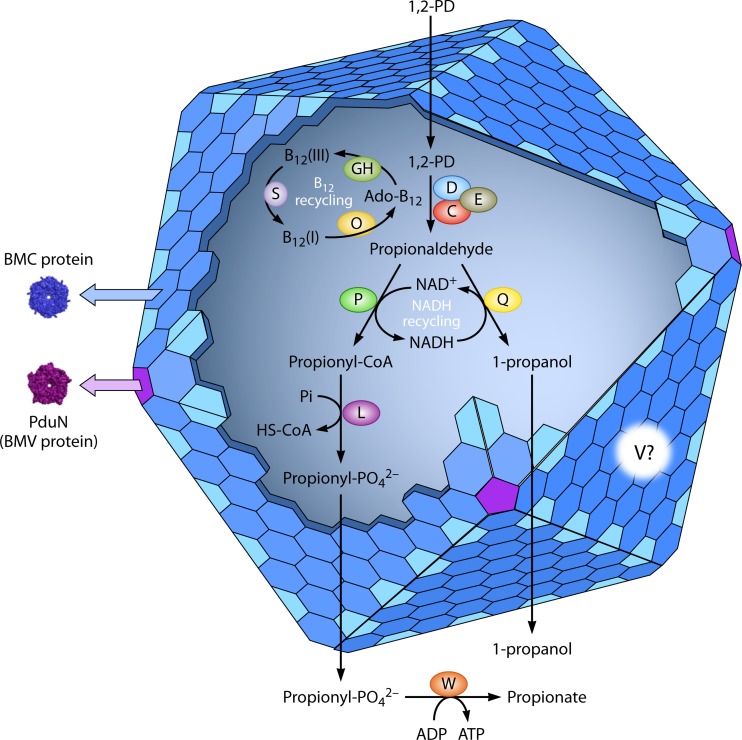
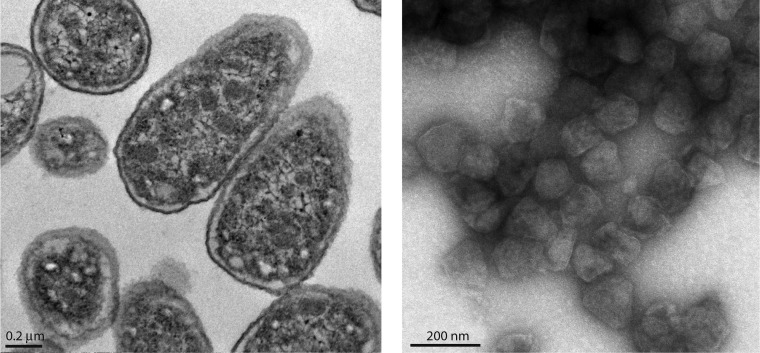
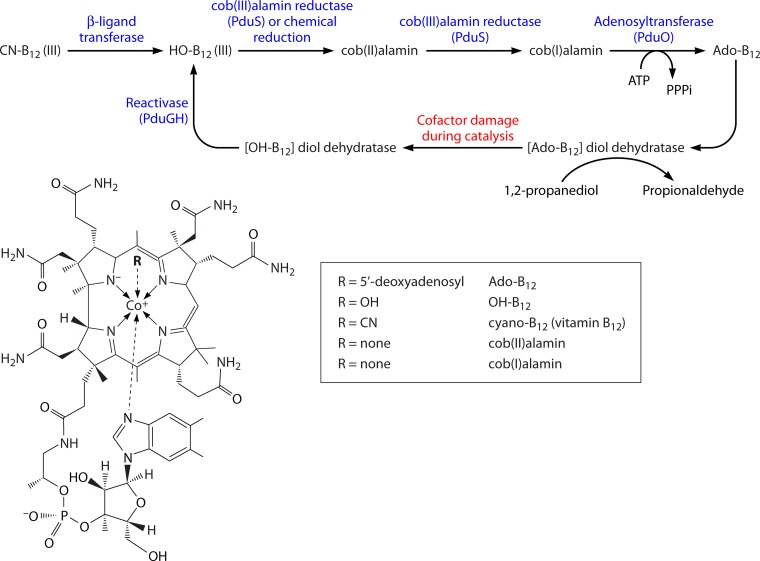
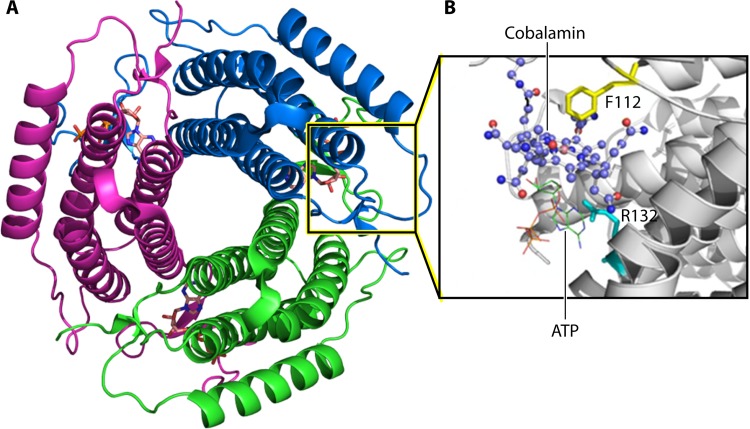
Bacterial microcompartments (MCPs) are sophisticated protein-based organelles used to optimize metabolic pathways. They consist of metabolic enzymes encapsulated within a protein shell, which creates an ideal environment for catalysis and facilitates the channeling of toxic/volatile intermediates to downstream enzymes. The metabolic processes that require MCPs are diverse and widely distributed and play important roles in global carbon fixation and bacterial pathogenesis. The protein shells of MCPs are thought to selectively control the movement of enzyme cofactors, substrates, and products (including toxic or volatile intermediates) between the MCP interior and the cytoplasm of the cell using both passive electrostatic/steric and dynamic gated mechanisms. Evidence suggests that specialized shell proteins conduct electrons between the cytoplasm and the lumen of the MCP and/or help rebuild damaged iron-sulfur centers in the encapsulated enzymes. The MCP shell is elaborated through a family of small proteins whose structural core is known as a bacterial microcompartment (BMC) domain. BMC domain proteins oligomerize into flat, hexagonally shaped tiles, which assemble into extended protein sheets that form the facets of the shell. Shape complementarity along the edges allows different types of BMC domain proteins to form mixed sheets, while sequence variation provides functional diversification. Recent studies have also revealed targeting sequences that mediate protein encapsulation within MCPs, scaffolding proteins that organize lumen enzymes and the use of private cofactor pools (NAD/H and coenzyme A [HS-CoA]) to facilitate cofactor homeostasis. Although much remains to be learned, our growing understanding of MCPs is providing a basis for bioengineering of protein-based containers for the production of chemicals/pharmaceuticals and for use as molecular delivery vehicles.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
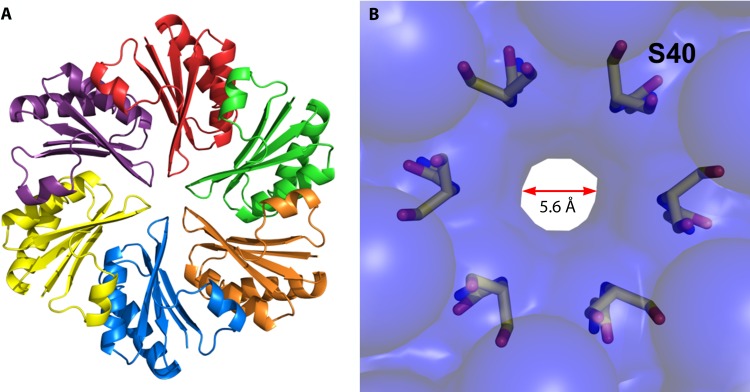
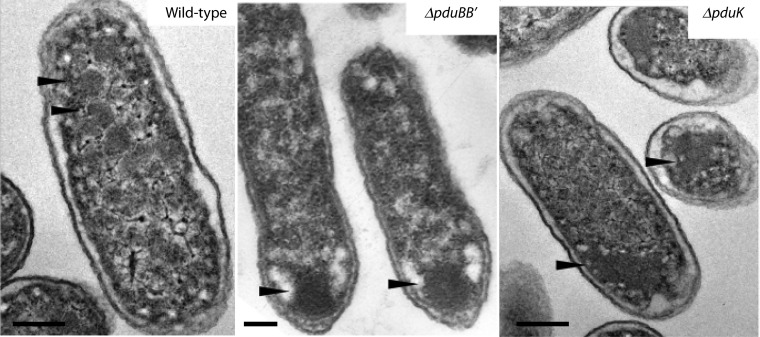
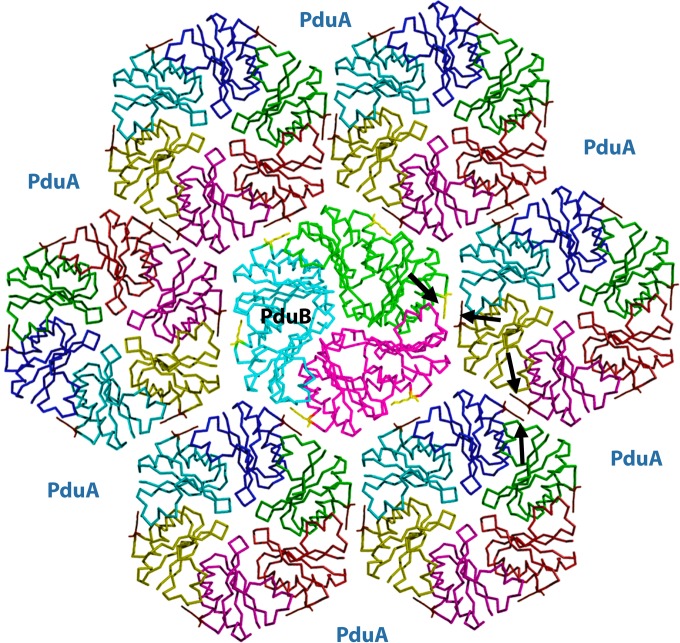
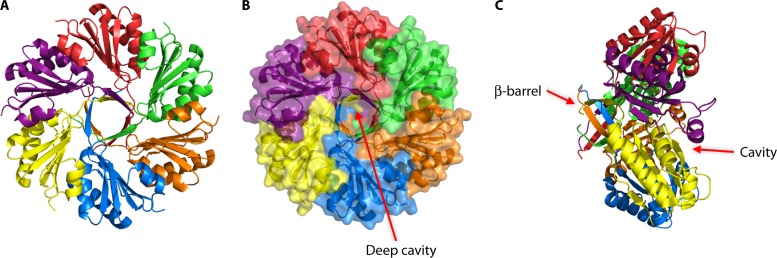
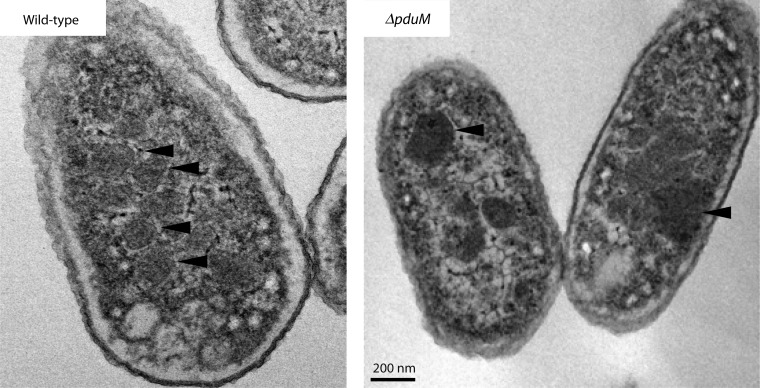
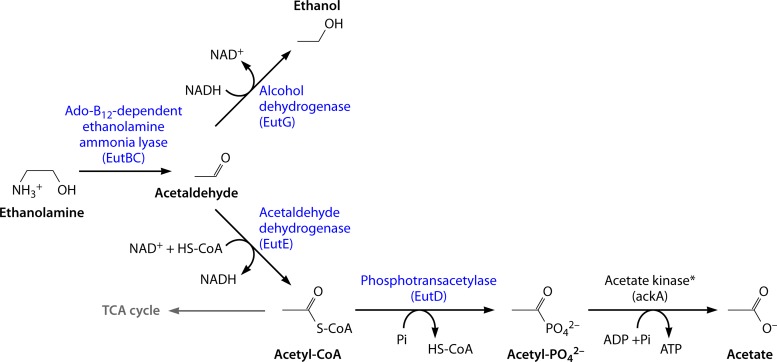
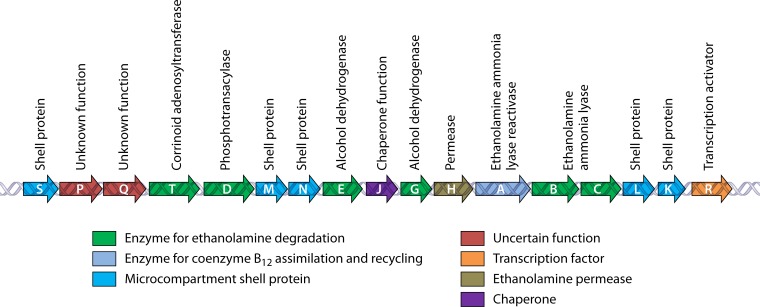
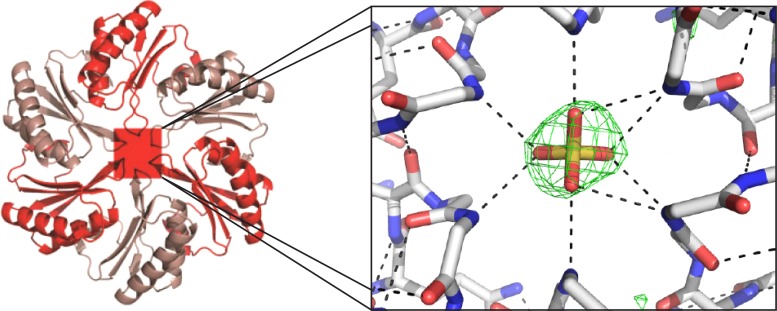
References
-
- Drews G, Niklowitz W. 1956. Beiträge zur Cytologie der Blaualgen. II. Zentroplasma und granulare Einschlüsse von Phormidium uncinatum. Arch. Mikrobiol. 24:147–162 - PubMed
-
- Cannon GC, Shively JM. 1983. Characterization of a homogenous preparation of carboxysomes from Thiobacillus neapolitanus. Arch. Microbiol. 134:52–59. 10.1007/BF00429407 - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
