

Search for microRNAs expressed by intracellular bacterial pathogens in infected mammalian cells
- PMID: 25184567
- PMCID: PMC4153649
- DOI: 10.1371/journal.pone.0106434
Search for microRNAs expressed by intracellular bacterial pathogens in infected mammalian cells
Abstract
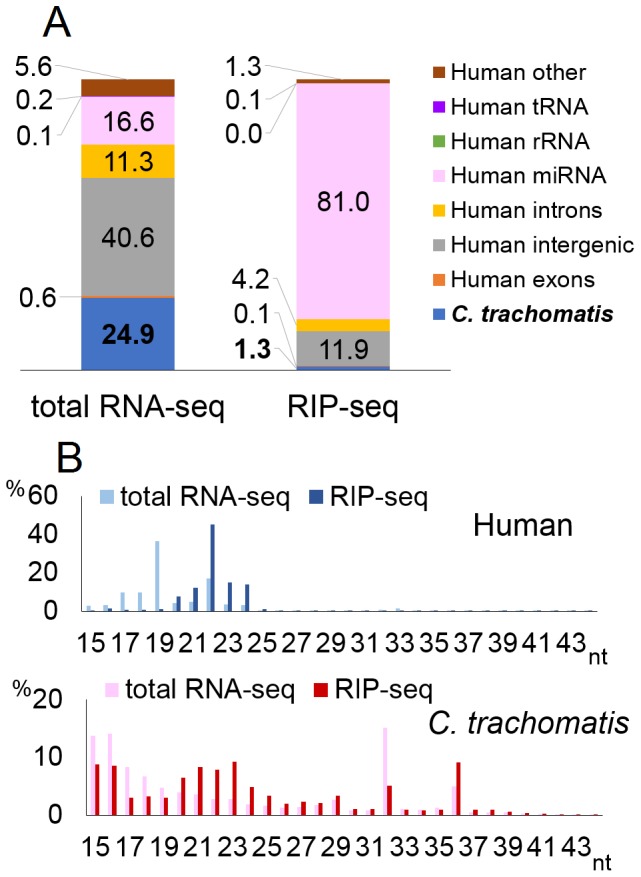
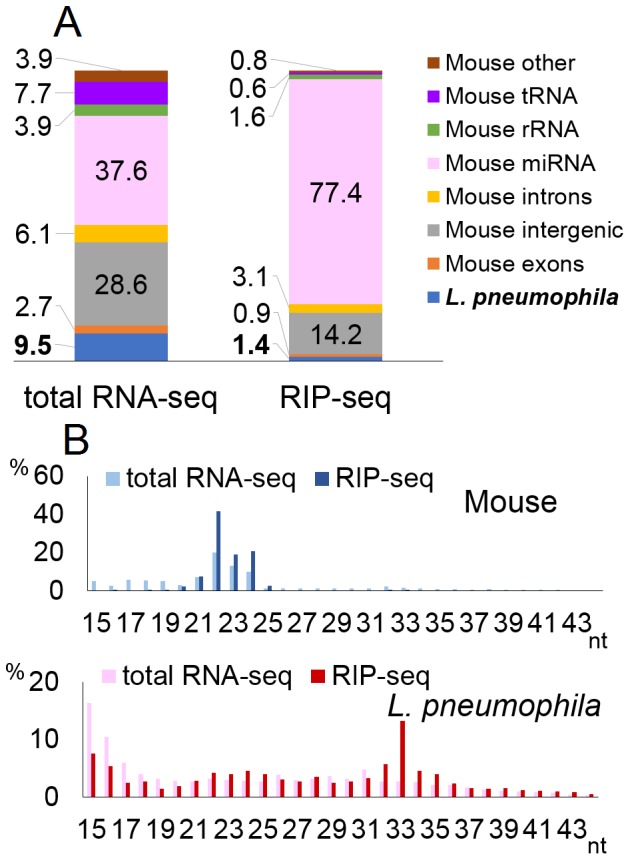
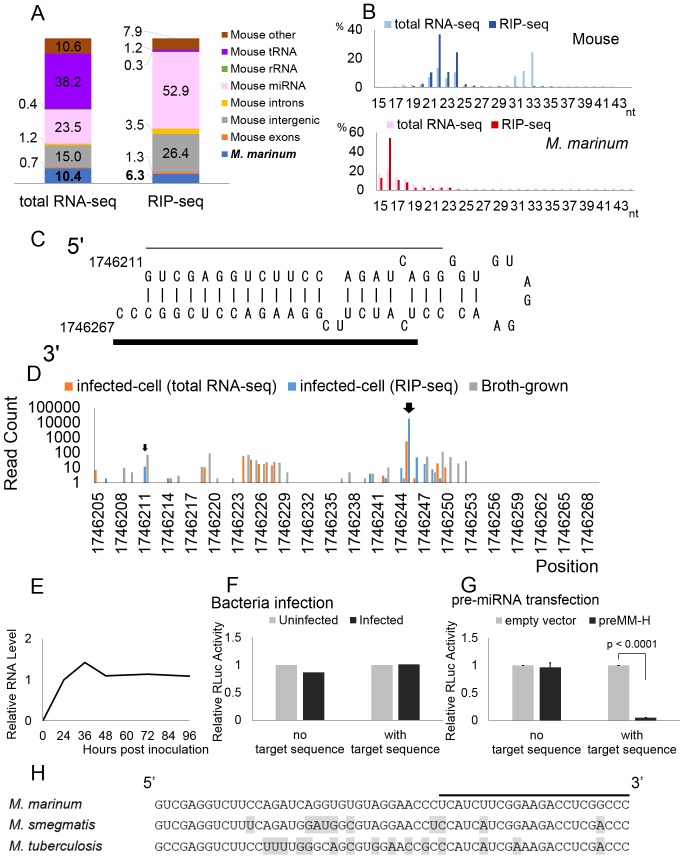
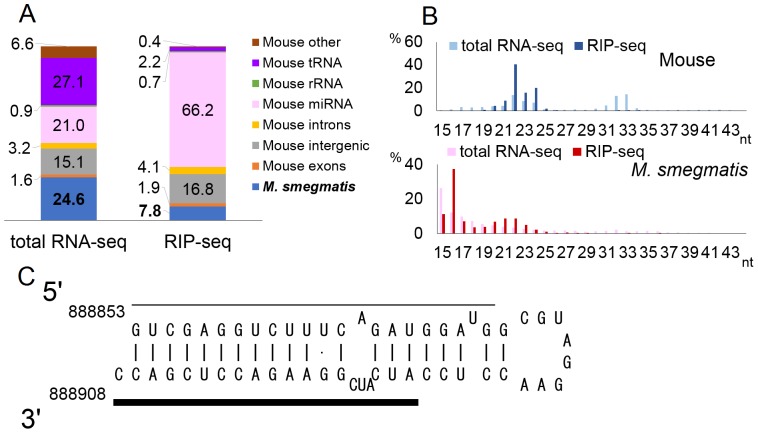
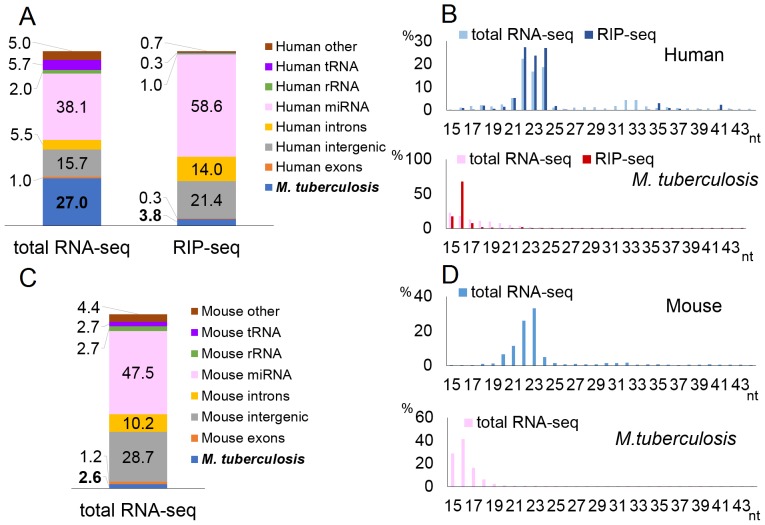
MicroRNAs are expressed by all multicellular organisms and play a critical role as post-transcriptional regulators of gene expression. Moreover, different microRNA species are known to influence the progression of a range of different diseases, including cancer and microbial infections. A number of different human viruses also encode microRNAs that can attenuate cellular innate immune responses and promote viral replication, and a fungal pathogen that infects plants has recently been shown to express microRNAs in infected cells that repress host cell immune responses and promote fungal pathogenesis. Here, we have used deep sequencing of total expressed small RNAs, as well as small RNAs associated with the cellular RNA-induced silencing complex RISC, to search for microRNAs that are potentially expressed by intracellular bacterial pathogens and translocated into infected animal cells. In the case of Legionella and Chlamydia and the two mycobacterial species M. smegmatis and M. tuberculosis, we failed to detect any bacterial small RNAs that had the characteristics expected for authentic microRNAs, although large numbers of small RNAs of bacterial origin could be recovered. However, a third mycobacterial species, M. marinum, did express an ∼ 23-nt small RNA that was bound by RISC and derived from an RNA stem-loop with the characteristics expected for a pre-microRNA. While intracellular expression of this candidate bacterial microRNA was too low to effectively repress target mRNA species in infected cultured cells in vitro, artificial overexpression of this potential bacterial pre-microRNA did result in the efficient repression of a target mRNA. This bacterial small RNA therefore represents the first candidate microRNA of bacterial origin.
Conflict of interest statement
Figures
References
-
- Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, et al. (1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 391: 806–811. - PubMed
-
- Martinez J, Patkaniowska A, Urlaub H, Luhrmann R, Tuschl T (2002) Single-stranded antisense siRNAs guide target RNA cleavage in RNAi. Cell 110: 563–574. - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ (2001) Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature 409: 363–366. - PubMed
-
- Hammond SM, Bernstein E, Beach D, Hannon GJ (2000) An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature 404: 293–296. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
