

Dynactin functions as both a dynamic tether and brake during dynein-driven motility
- PMID: 25185702
- PMCID: PMC4470572
- DOI: 10.1038/ncomms5807
Dynactin functions as both a dynamic tether and brake during dynein-driven motility
Abstract
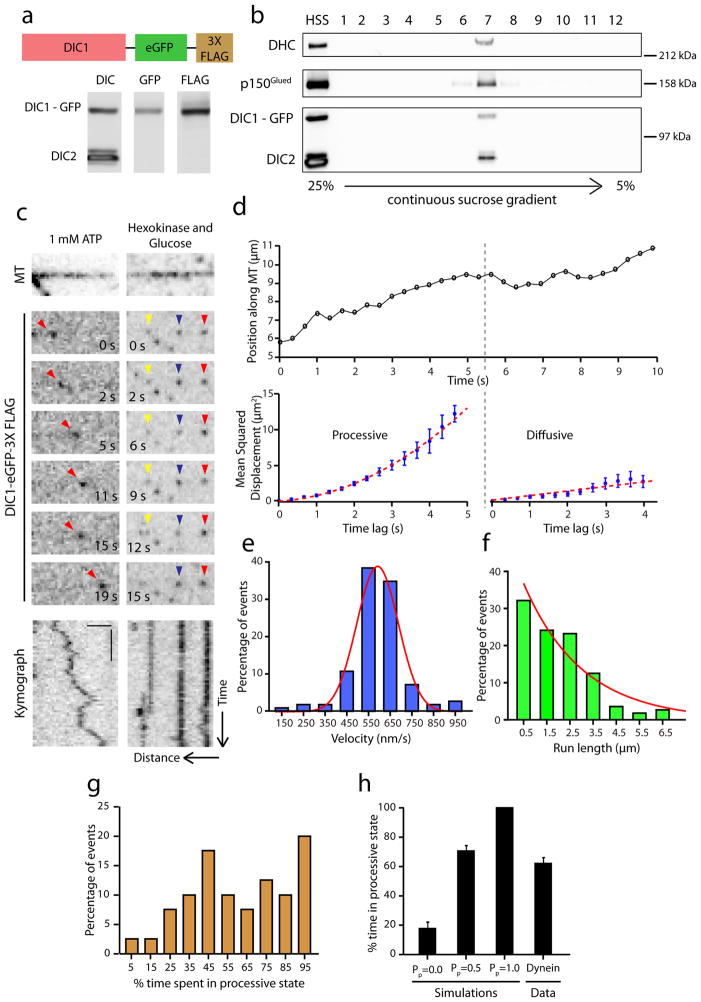
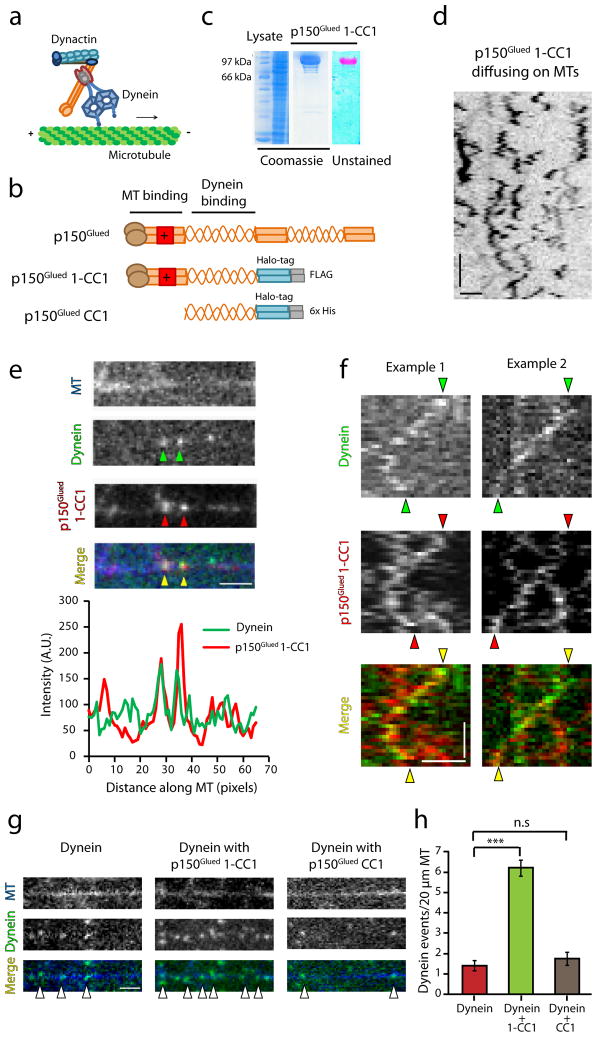
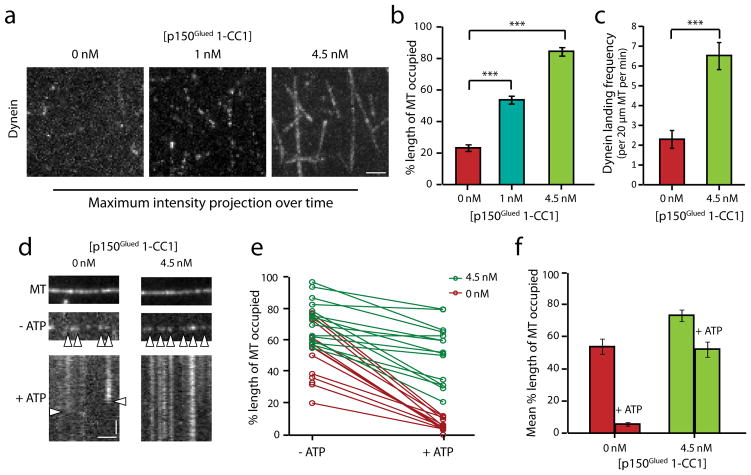
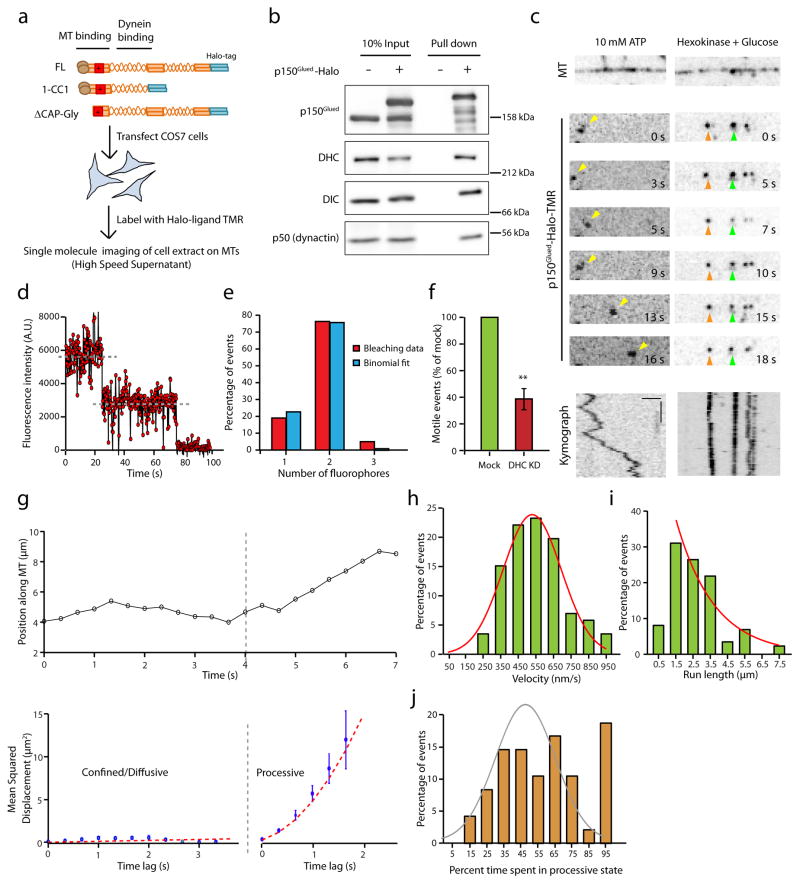
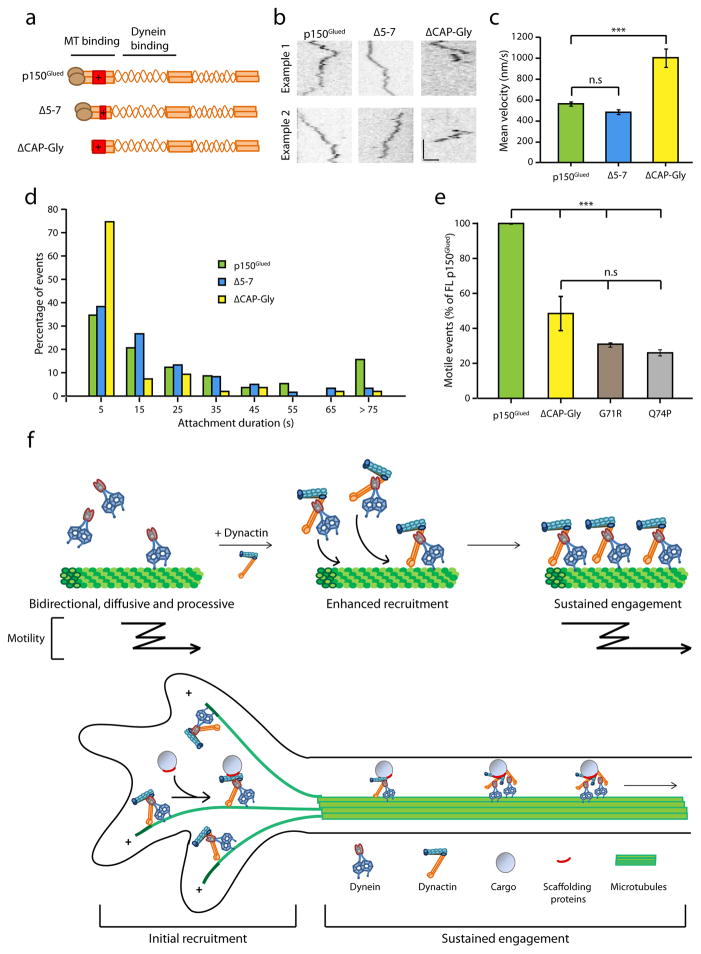
Dynactin is an essential cofactor for most cellular functions of the microtubule motor cytoplasmic dynein, but the mechanism by which dynactin activates dynein remains unclear. Here we use single molecule approaches to investigate dynein regulation by the dynactin subunit p150(Glued). We investigate the formation and motility of a dynein-p150(Glued) co-complex using dual-colour total internal reflection fluorescence microscopy. p150(Glued) recruits and tethers dynein to the microtubule in a concentration-dependent manner. Single molecule imaging of motility in cell extracts demonstrates that the CAP-Gly domain of p150(Glued) decreases the detachment rate of the dynein-dynactin complex from the microtubule and also acts as a brake to slow the dynein motor. Consistent with this important role, two neurodegenerative disease-causing mutations in the CAP-Gly domain abrogate these functions in our assays. Together, these observations support a model in which dynactin enhances the initial recruitment of dynein onto microtubules and promotes the sustained engagement of dynein with its cytoskeletal track.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Schroer TA. Dynactin. Annu Rev Cell Dev Biol. 2004;20:759–79. - PubMed
-
- Holzbaur ELF, et al. Homology of a 150K cytoplasmic dynein- associated polypeptide with the Drosophila gene Glued. Nature. 1991;351:579–583. - PubMed
-
- Karki S, Holzbaur ELF. Affinity Chromatography Demonstrates a Direct Binding between Cytoplasmic Dynein and the Dynactin Complex. J Biol Chem. 1995;270:28806–28811. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
