

Mitogen-activated protein kinase-interacting kinase regulates mTOR/AKT signaling and controls the serine/arginine-rich protein kinase-responsive type 1 internal ribosome entry site-mediated translation and viral oncolysis
- PMID: 25187540
- PMCID: PMC4249080
- DOI: 10.1128/JVI.01884-14
Mitogen-activated protein kinase-interacting kinase regulates mTOR/AKT signaling and controls the serine/arginine-rich protein kinase-responsive type 1 internal ribosome entry site-mediated translation and viral oncolysis
Abstract
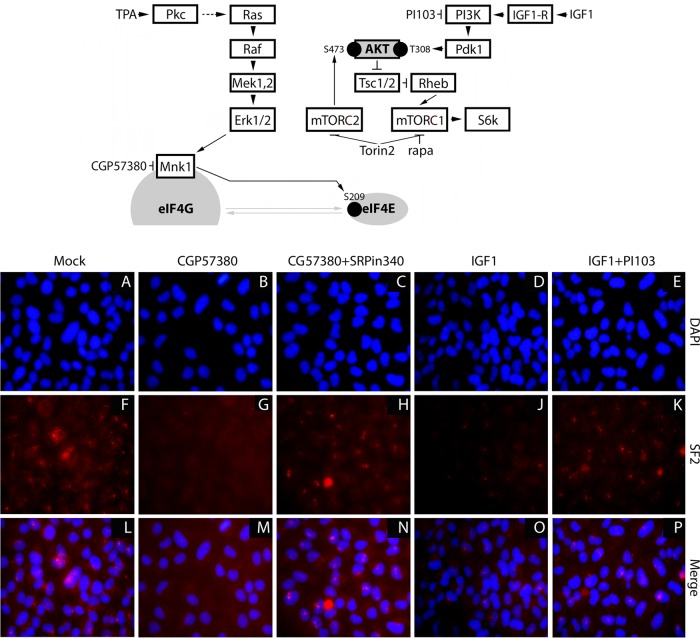
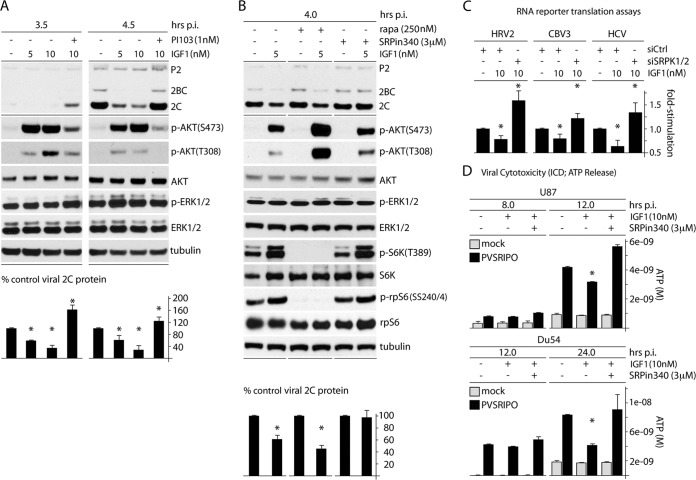
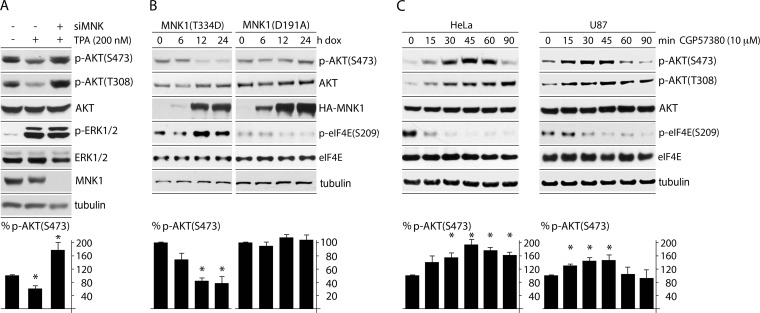
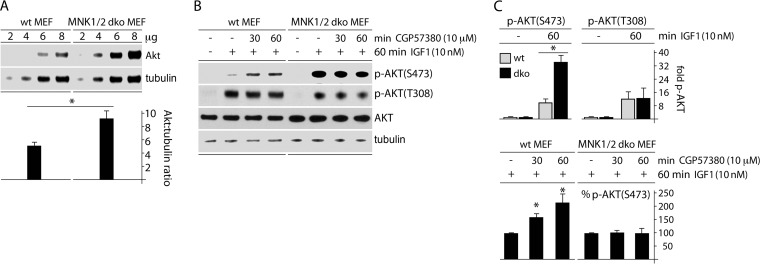
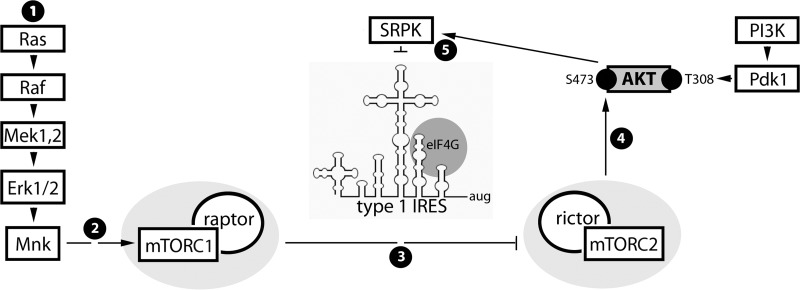
Translation machinery is a major recipient of the principal mitogenic signaling networks involving Raf-ERK1/2 and phosphoinositol 3-kinase (PI3K)-mechanistic target of rapamycin (mTOR). Picornavirus internal ribosomal entry site (IRES)-mediated translation and cytopathogenic effects are susceptible to the status of such signaling cascades in host cells. We determined that tumor-specific cytotoxicity of the poliovirus/rhinovirus chimera PVSRIPO is facilitated by Raf-ERK1/2 signals to the mitogen-activated protein kinase (MAPK)-interacting kinase (MNK) and its effects on the partitioning/activity of the Ser/Arg (SR)-rich protein kinase (SRPK) (M. C. Brown, J. D. Bryant, E. Y. Dobrikova, M. Shveygert, S. S. Bradrick, V. Chandramohan, D. D. Bigner, and M, Gromeier, J. Virol. 22:13135-13148, 2014, doi:http://dx.doi.org/10.1128/JVI.01883-14). Here, we show that MNK regulates SRPK via mTOR and AKT. Our investigations revealed a MNK-controlled mechanism acting on mTORC2-AKT. The resulting suppression of AKT signaling attenuates SRPK activity to enhance picornavirus type 1 IRES translation and favor PVSRIPO tumor cell toxicity and killing.
Importance: Oncolytic immunotherapy with PVSRIPO, the type 1 live-attenuated poliovirus (PV) (Sabin) vaccine containing a human rhinovirus type 2 (HRV2) IRES, is demonstrating early promise in clinical trials with intratumoral infusion in recurrent glioblastoma (GBM). Our investigations demonstrate that the core mechanistic principle of PVSRIPO, tumor-selective translation and cytotoxicity, relies on constitutive ERK1/2-MNK signals that counteract the deleterious effects of runaway AKT-SRPK activity in malignancy.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
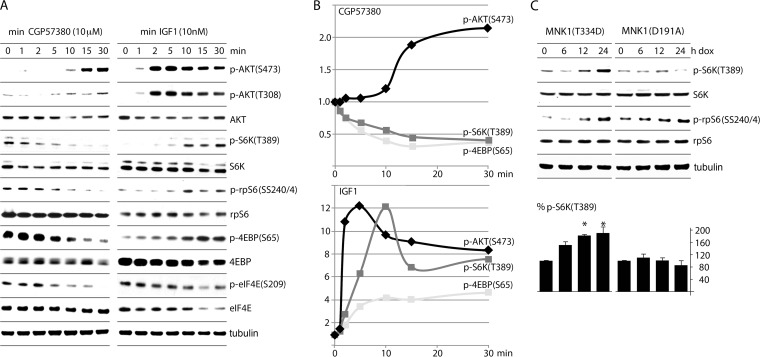
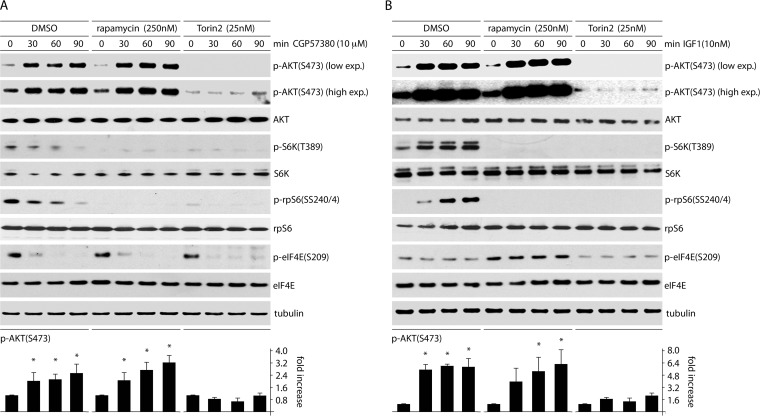
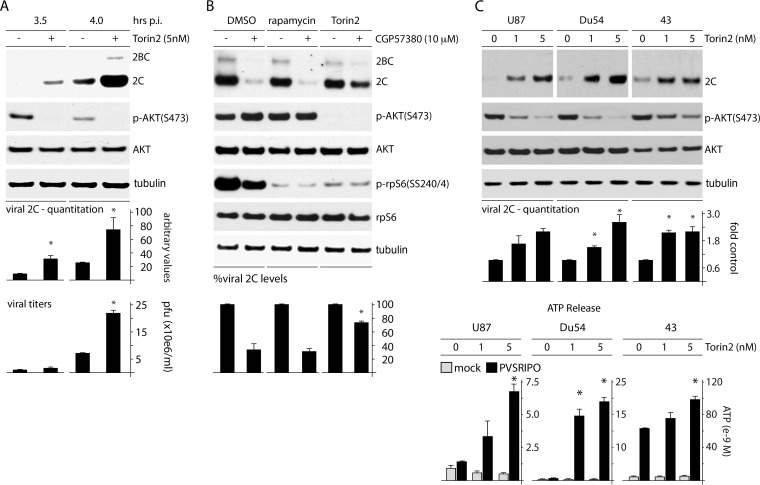
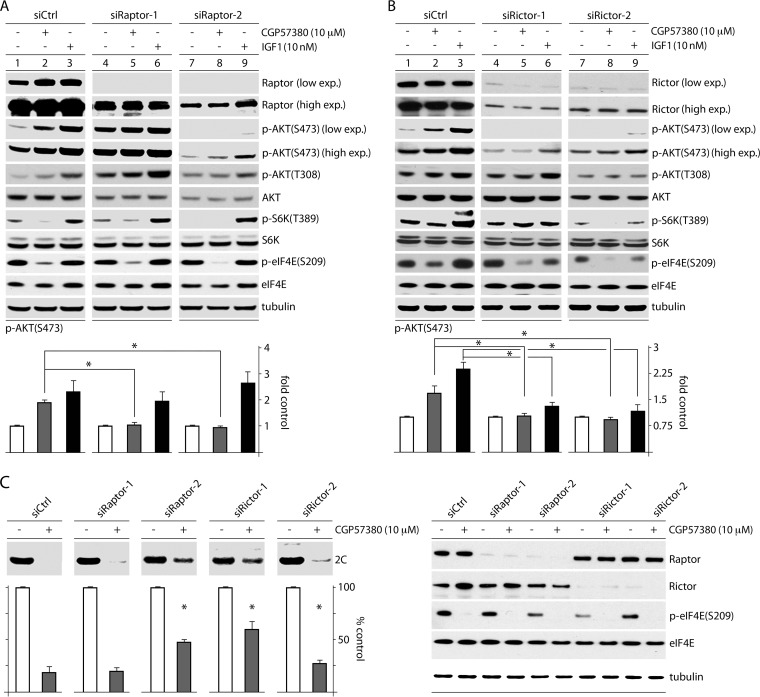
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
