

Induction of viral, 7-methyl-guanosine cap-independent translation and oncolysis by mitogen-activated protein kinase-interacting kinase-mediated effects on the serine/arginine-rich protein kinase
- PMID: 25187541
- PMCID: PMC4249076
- DOI: 10.1128/JVI.01883-14
Induction of viral, 7-methyl-guanosine cap-independent translation and oncolysis by mitogen-activated protein kinase-interacting kinase-mediated effects on the serine/arginine-rich protein kinase
Abstract
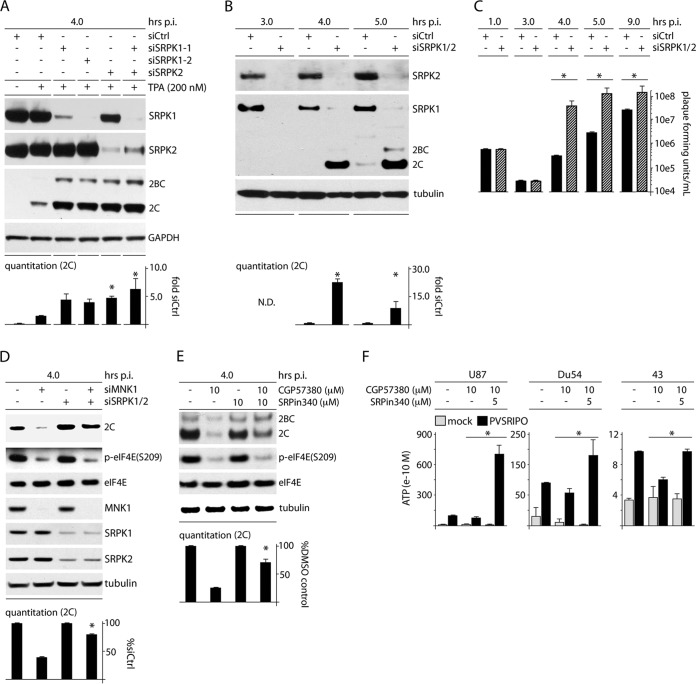
Protein synthesis, the most energy-consuming process in cells, responds to changing physiologic priorities, e.g., upon mitogen- or stress-induced adaptations signaled through the mitogen-activated protein kinases (MAPKs). The prevailing status of protein synthesis machinery is a viral pathogenesis factor, particularly for plus-strand RNA viruses, where immediate translation of incoming viral RNAs shapes host-virus interactions. In this study, we unraveled signaling pathways centered on the ERK1/2 and p38α MAPK-interacting kinases MNK1/2 and their role in controlling 7-methyl-guanosine (m(7)G) "cap"-independent translation at enterovirus type 1 internal ribosomal entry sites (IRESs). Activation of Raf-MEK-ERK1/2 signals induced viral IRES-mediated translation in a manner dependent on MNK1/2. This effect was not due to MNK's known functions as eukaryotic initiation factor (eIF) 4G binding partner or eIF4E(S209) kinase. Rather, MNK catalytic activity enabled viral IRES-mediated translation/host cell cytotoxicity through negative regulation of the Ser/Arg (SR)-rich protein kinase (SRPK). Our investigations suggest that SRPK activity is a major determinant of type 1 IRES competency, host cell cytotoxicity, and viral proliferation in infected cells.
Importance: We are targeting unfettered enterovirus IRES activity in cancer with PVSRIPO, the type 1 live-attenuated poliovirus (PV) (Sabin) vaccine containing a human rhinovirus type 2 (HRV2) IRES. A phase I clinical trial of PVSRIPO with intratumoral inoculation in patients with recurrent glioblastoma (GBM) is showing early promise. Viral translation proficiency in infected GBM cells is a core requirement for the antineoplastic efficacy of PVSRIPO. Therefore, it is critically important to understand the mechanisms controlling viral cap-independent translation in infected host cells.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
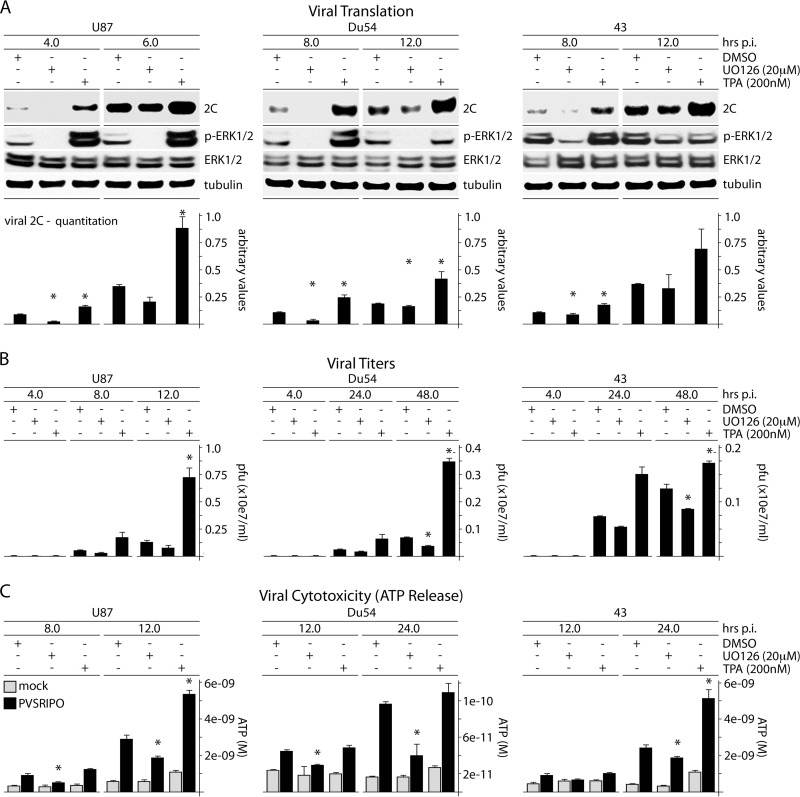
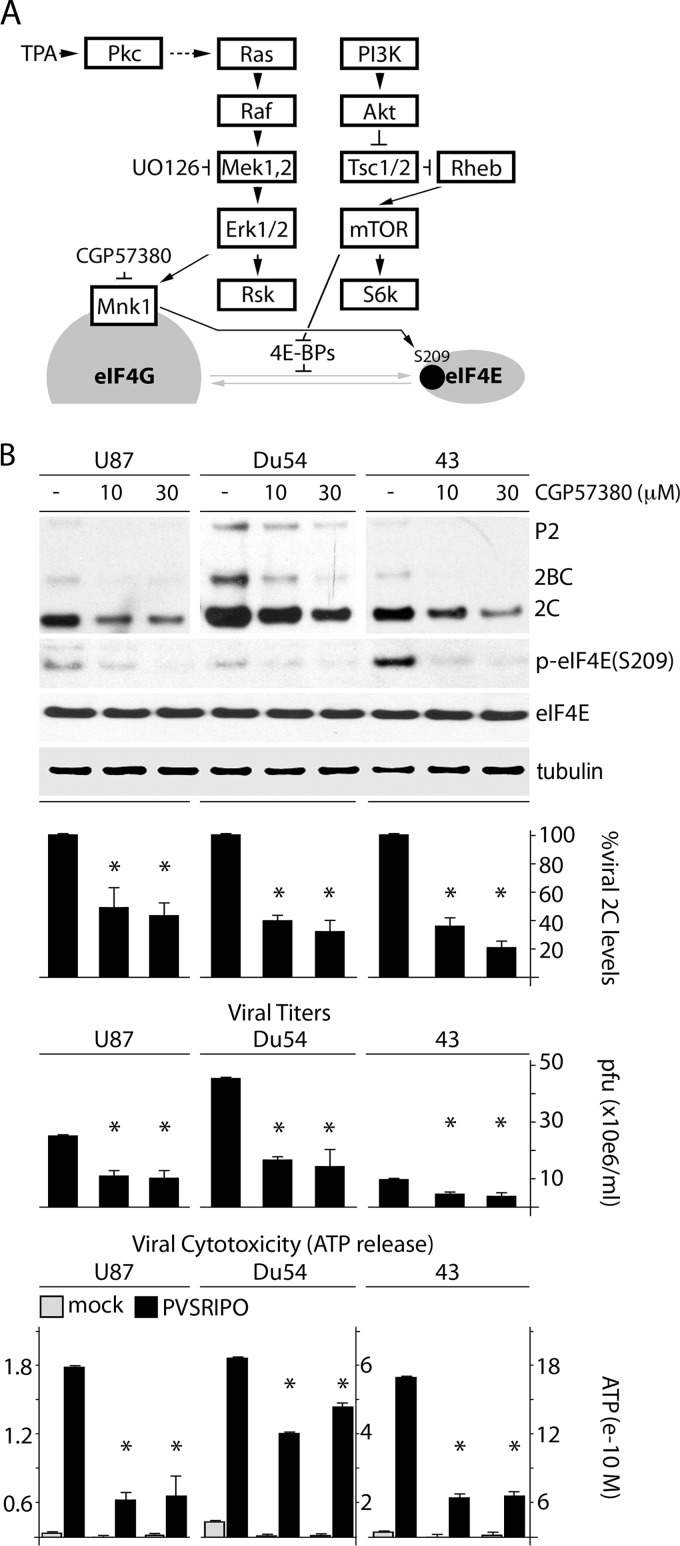
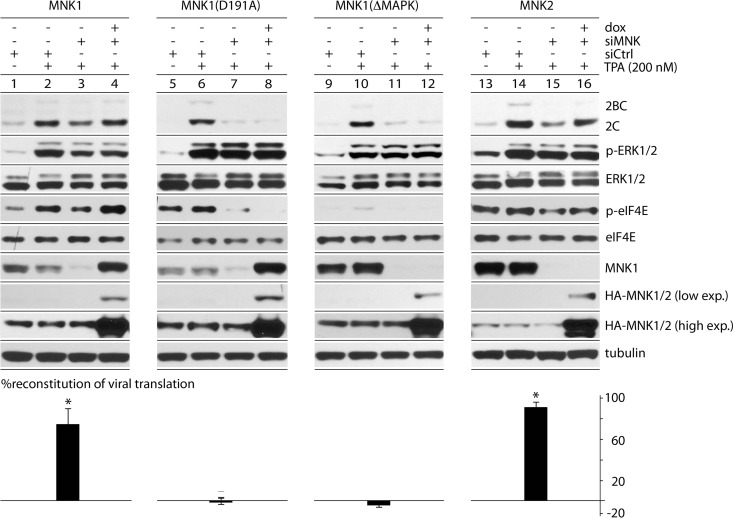
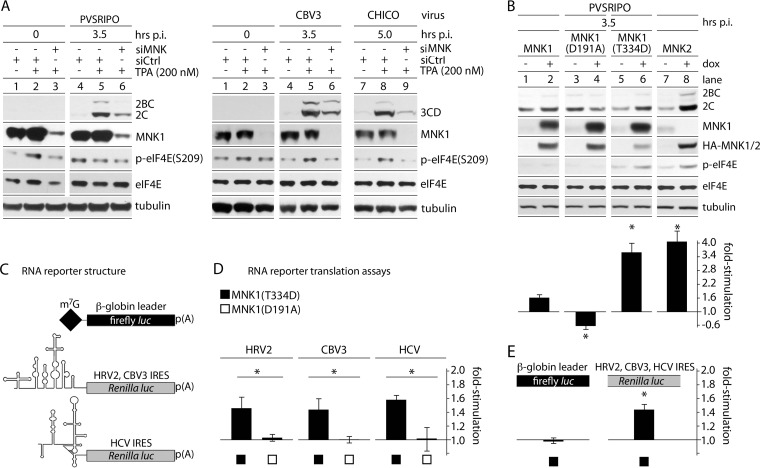
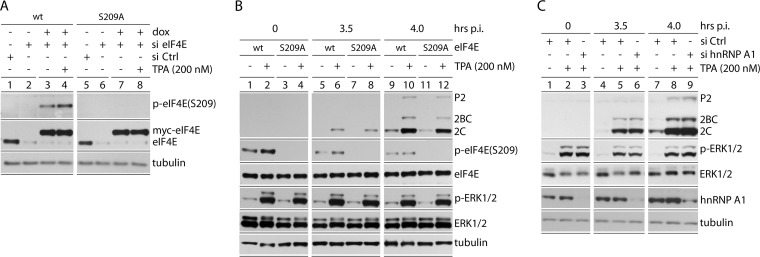
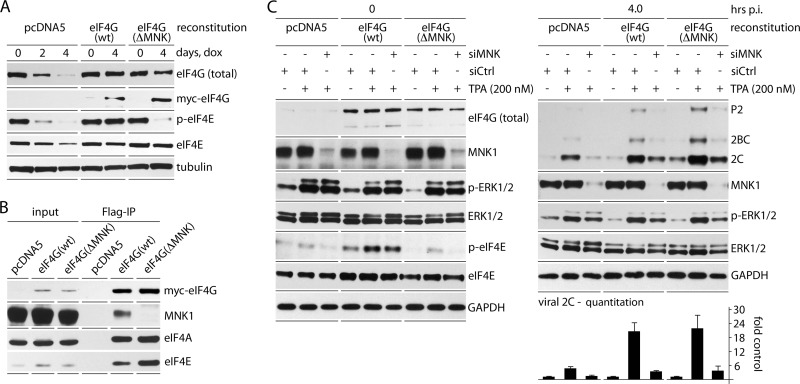
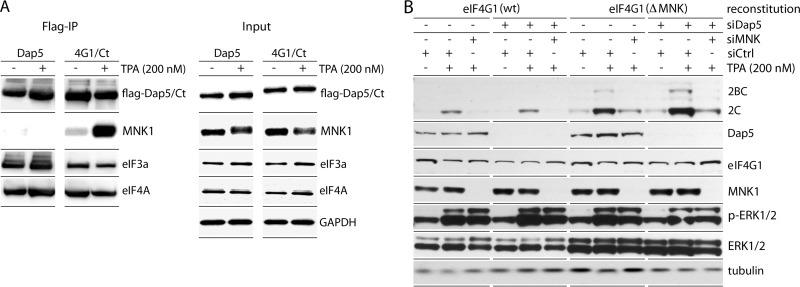
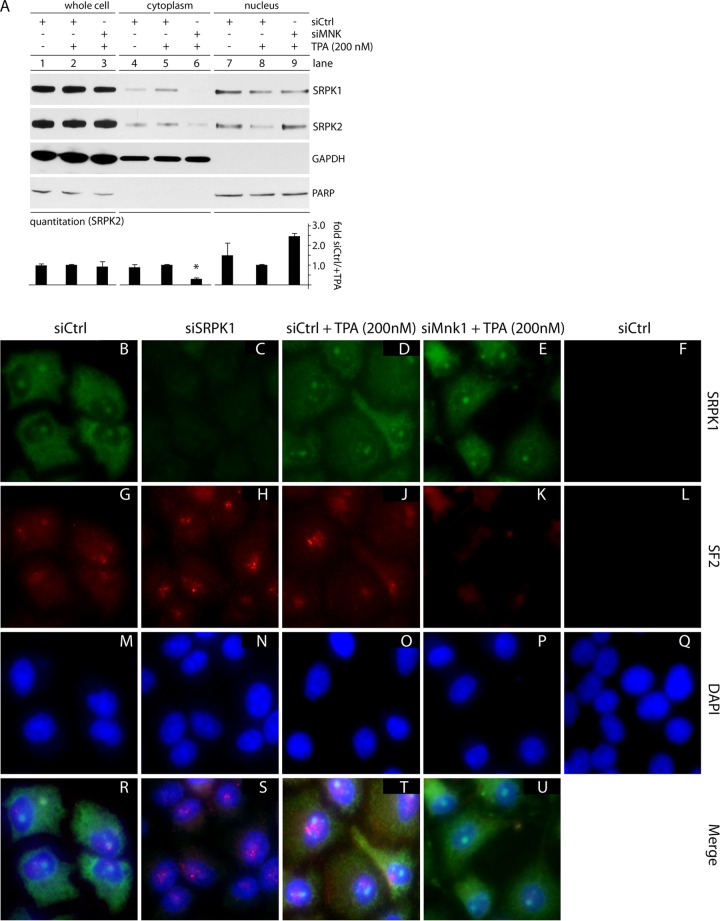
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
