

The olfactory transcriptomes of mice
- PMID: 25187969
- PMCID: PMC4154679
- DOI: 10.1371/journal.pgen.1004593
The olfactory transcriptomes of mice
Abstract
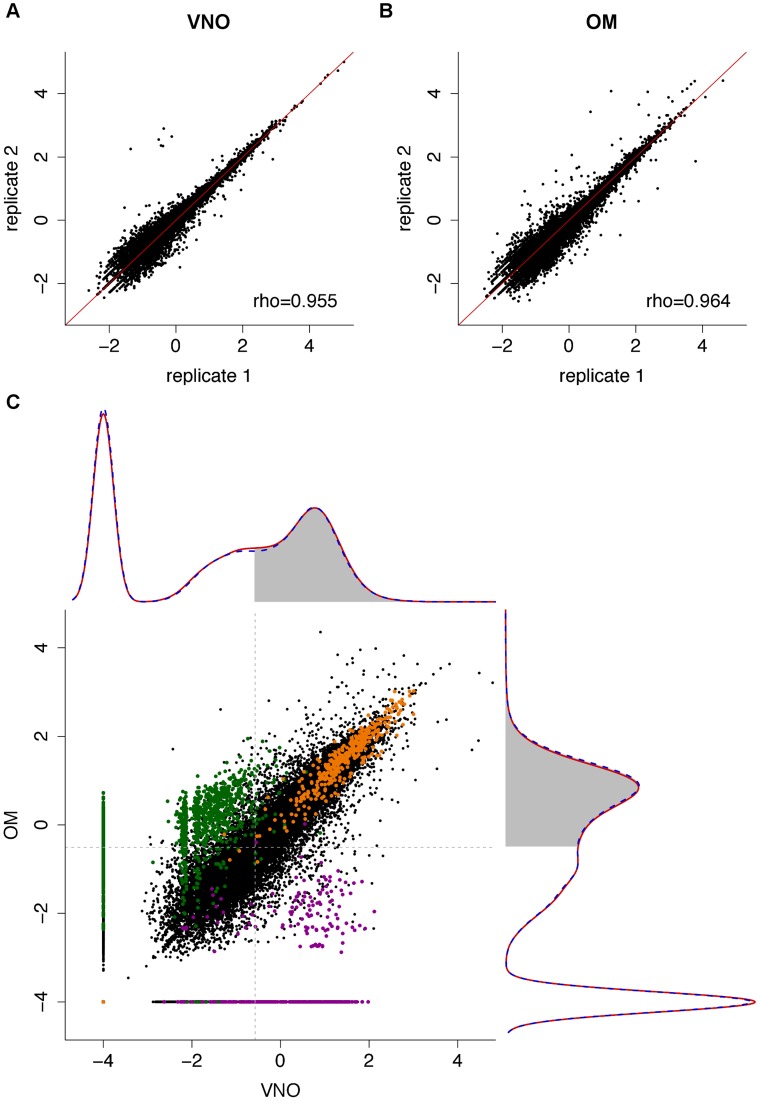
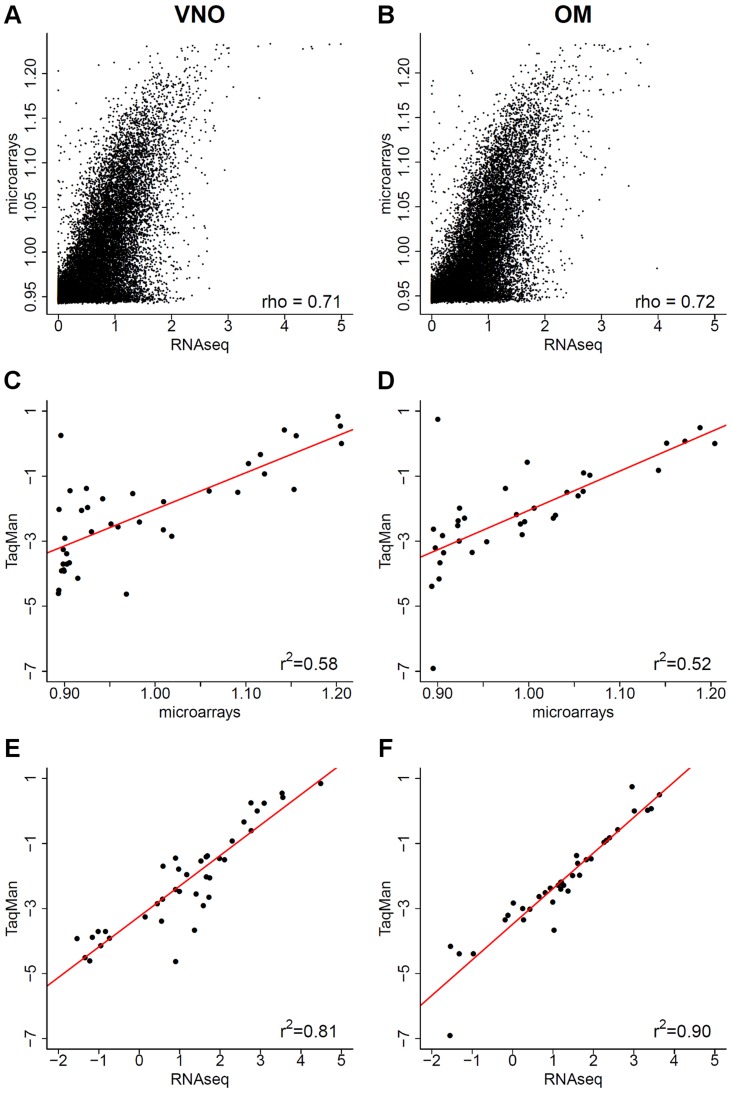
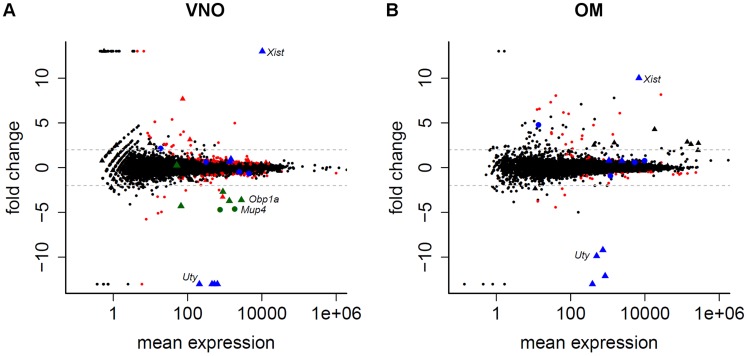
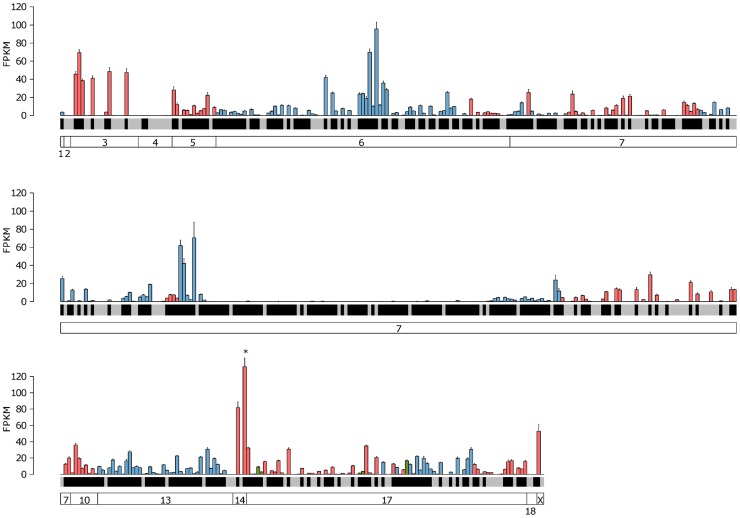
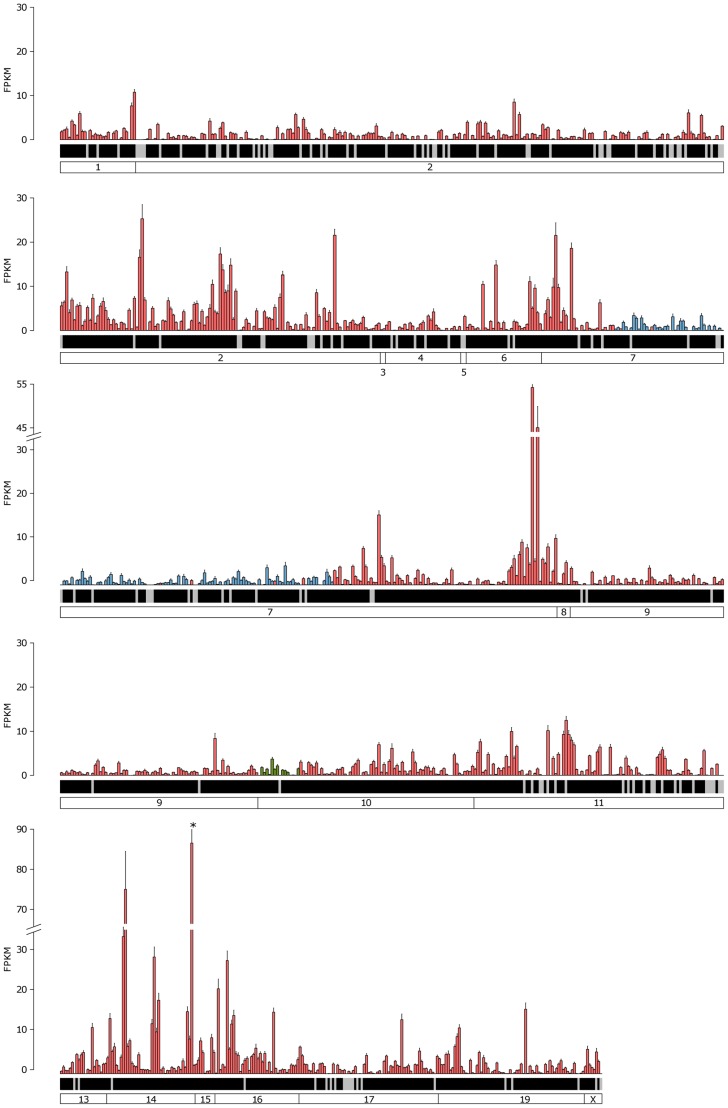
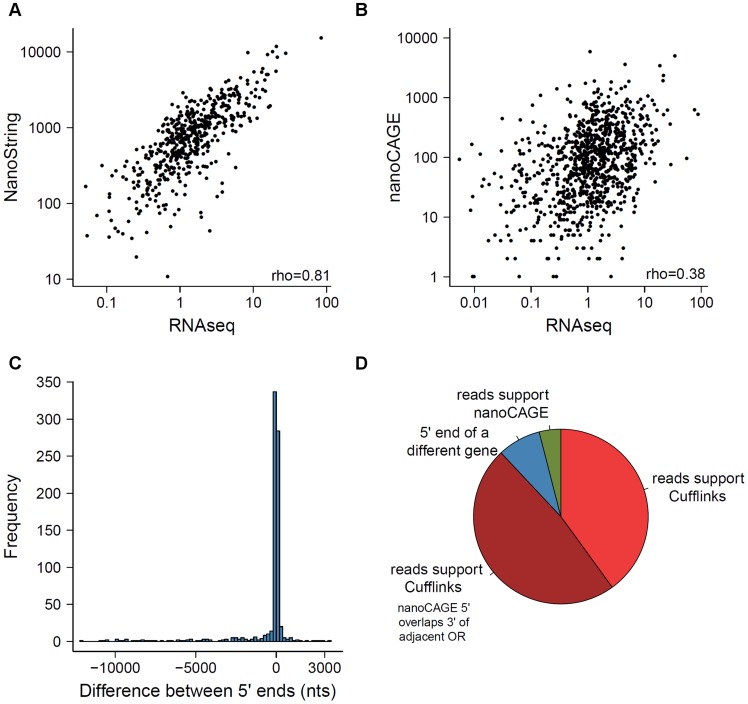
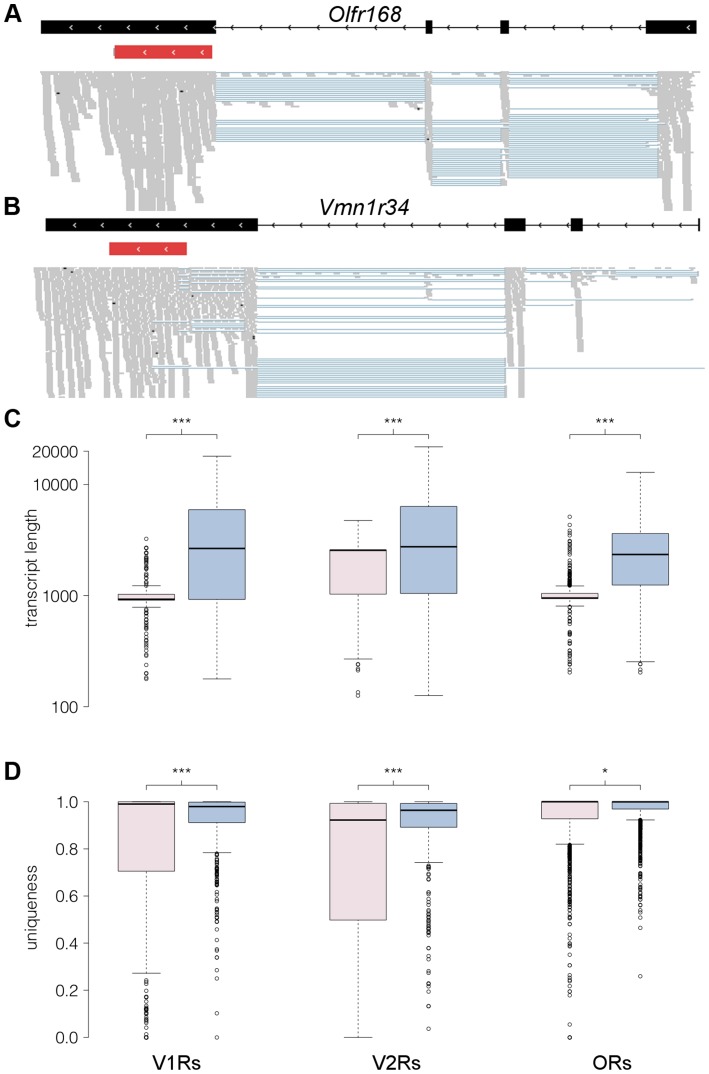
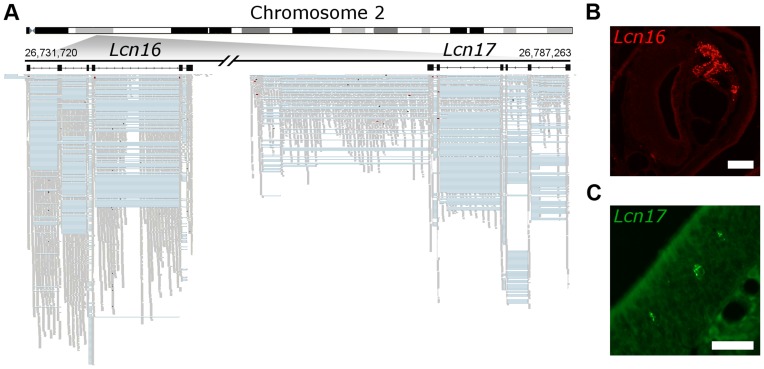
The olfactory (OR) and vomeronasal receptor (VR) repertoires are collectively encoded by 1700 genes and pseudogenes in the mouse genome. Most OR and VR genes were identified by comparative genomic techniques and therefore, in many of those cases, only their protein coding sequences are defined. Some also lack experimental support, due in part to the similarity between them and their monogenic, cell-specific expression in olfactory tissues. Here we use deep RNA sequencing, expression microarray and quantitative RT-PCR in both the vomeronasal organ and whole olfactory mucosa to quantify their full transcriptomes in multiple male and female mice. We find evidence of expression for all VR, and almost all OR genes that are annotated as functional in the reference genome, and use the data to generate over 1100 new, multi-exonic, significantly extended receptor gene annotations. We find that OR and VR genes are neither equally nor randomly expressed, but have reproducible distributions of abundance in both tissues. The olfactory transcriptomes are only minimally different between males and females, suggesting altered gene expression at the periphery is unlikely to underpin the striking sexual dimorphism in olfactory-mediated behavior. Finally, we present evidence that hundreds of novel, putatively protein-coding genes are expressed in these highly specialized olfactory tissues, and carry out a proof-of-principle validation. Taken together, these data provide a comprehensive, quantitative catalog of the genes that mediate olfactory perception and pheromone-evoked behavior at the periphery.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Dulac C, Torello AT (2003) Molecular detection of pheromone signals in mammals: from genes to behaviour. Nat Rev Neurosci 4: 551–562. - PubMed
-
- Chamero P, Marton TF, Logan DW, Flanagan K, Cruz JR, et al. (2007) Identification of protein pheromones that promote aggressive behaviour. Nature 450: 899–902. - PubMed
-
- Haga S, Hattori T, Sato T, Sato K, Matsuda S, et al. (2010) The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature 466: 118–122. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
