

Eliminating both canonical and short-patch mismatch repair in Drosophila melanogaster suggests a new meiotic recombination model
- PMID: 25188408
- PMCID: PMC4154643
- DOI: 10.1371/journal.pgen.1004583
Eliminating both canonical and short-patch mismatch repair in Drosophila melanogaster suggests a new meiotic recombination model
Abstract
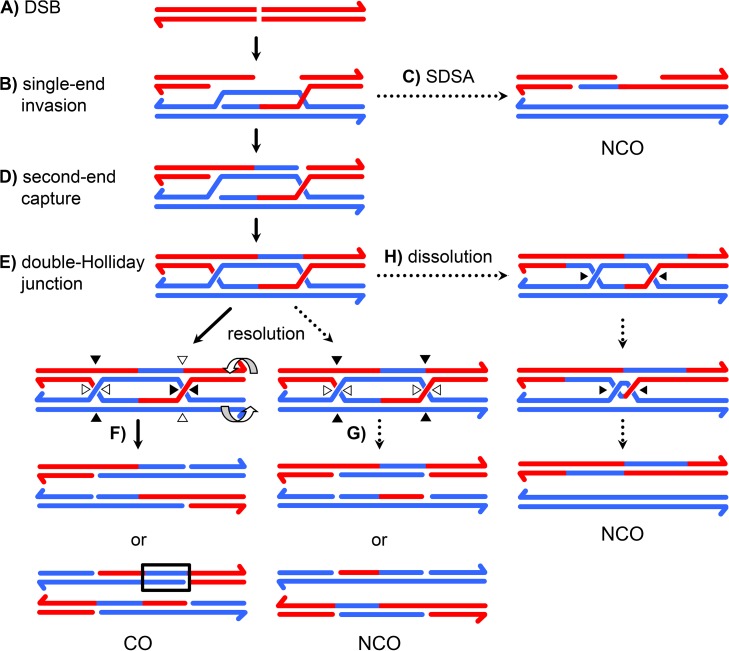
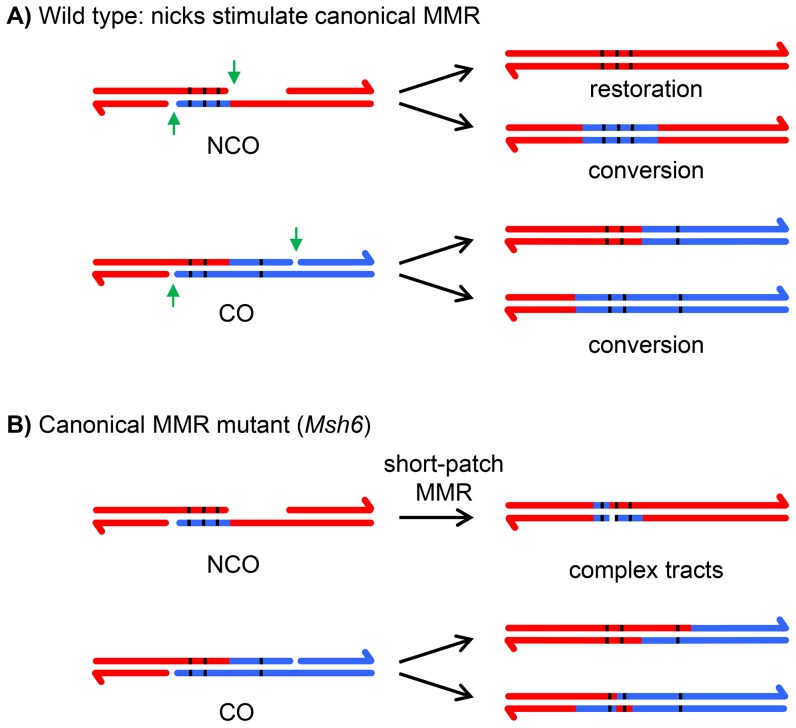
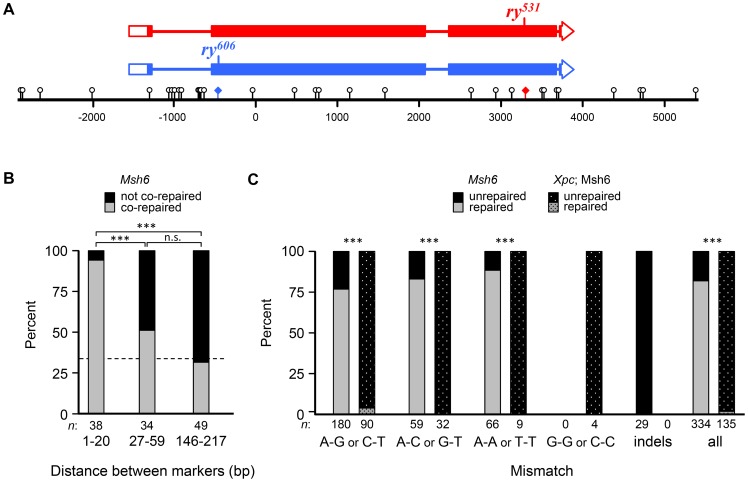
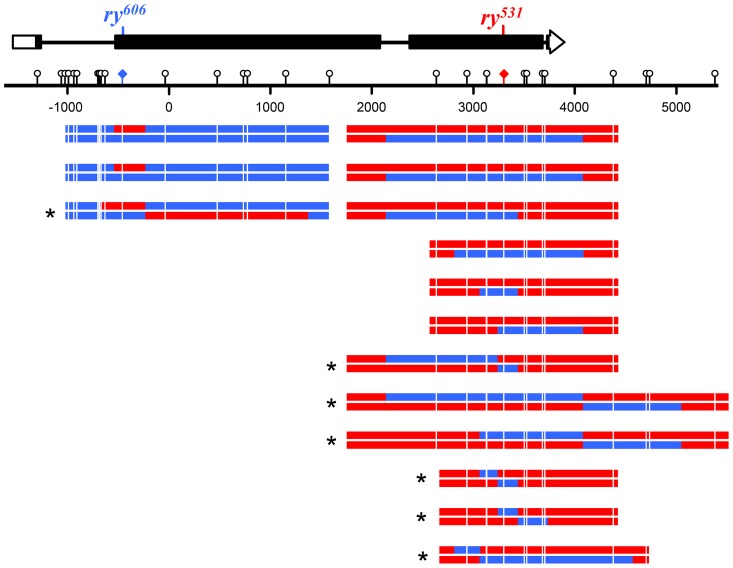
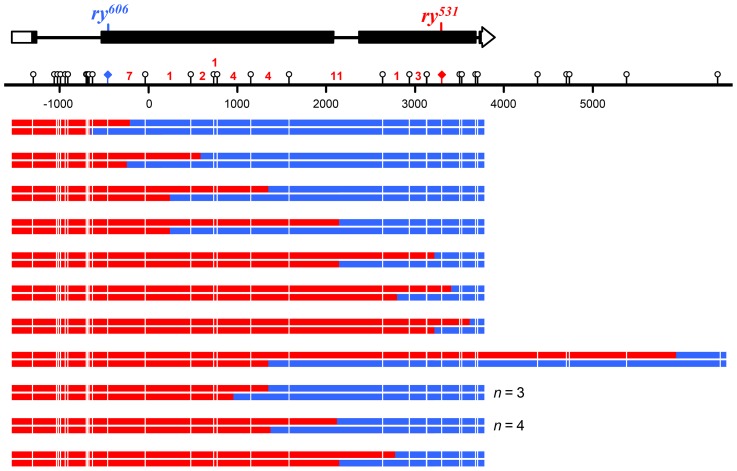
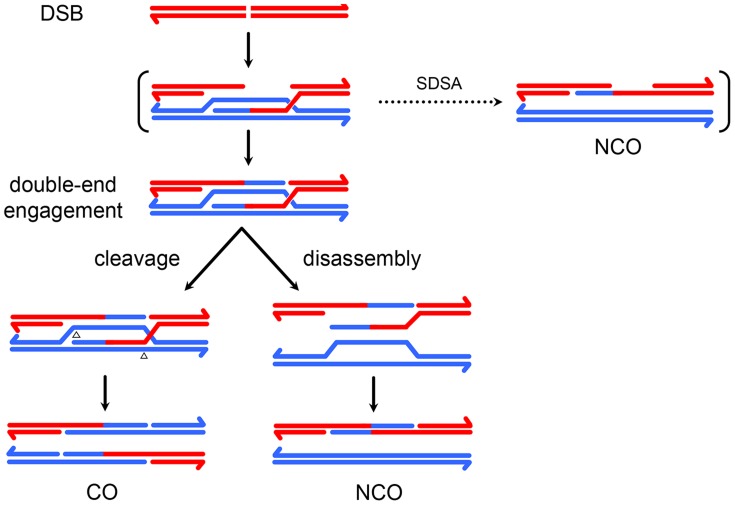
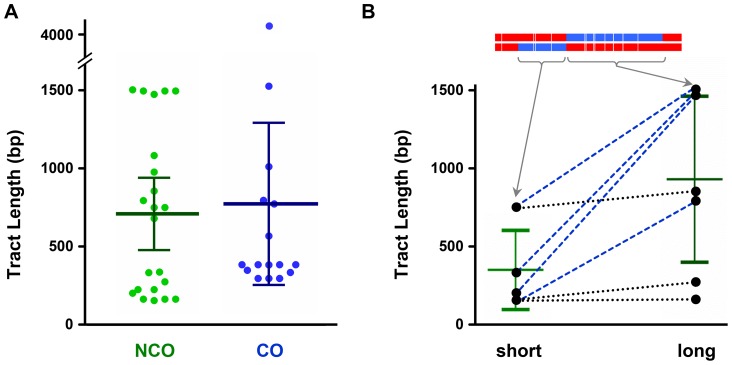
In most meiotic systems, recombination is essential to form connections between homologs that ensure their accurate segregation from one another. Meiotic recombination is initiated by DNA double-strand breaks that are repaired using the homologous chromosome as a template. Studies of recombination in budding yeast have led to a model in which most early repair intermediates are disassembled to produce noncrossovers. Selected repair events are stabilized so they can proceed to form double-Holliday junction (dHJ) intermediates, which are subsequently resolved into crossovers. This model is supported in yeast by physical isolation of recombination intermediates, but the extent to which it pertains to animals is unknown. We sought to test this model in Drosophila melanogaster by analyzing patterns of heteroduplex DNA (hDNA) in recombination products. Previous attempts to do this have relied on knocking out the canonical mismatch repair (MMR) pathway, but in both yeast and Drosophila the resulting recombination products are complex and difficult to interpret. We show that, in Drosophila, this complexity results from a secondary, short-patch MMR pathway that requires nucleotide excision repair. Knocking out both canonical and short-patch MMR reveals hDNA patterns that reveal that many noncrossovers arise after both ends of the break have engaged with the homolog. Patterns of hDNA in crossovers could be explained by biased resolution of a dHJ; however, considering the noncrossover and crossover results together suggests a model in which a two-end engagement intermediate with unligated HJs can be disassembled by a helicase to a produce noncrossover or nicked by a nuclease to produce a crossover. While some aspects of this model are similar to the model from budding yeast, production of both noncrossovers and crossovers from a single, late intermediate is a fundamental difference that has important implications for crossover control.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Bishop DK, Zickler D (2004) Early decision; meiotic crossover interference prior to stable strand exchange and synapsis. Cell 117: 9–15. - PubMed
-
- Schwacha A, Kleckner N (1995) Identification of double Holliday junctions as intermediates in meiotic recombination. Cell 83: 783–791. - PubMed
-
- Hunter N, Kleckner N (2001) The single-end invasion: an asymmetric intermediate at the double-strand break to double-holliday junction transition of meiotic recombination. Cell 106: 59–70. - PubMed
-
- Szostak JW, Orr-Weaver TL, Rothstein RJ, Stahl FW (1983) The double-strand-break repair model for recombination. Cell 33: 25–35. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
