

Olfactory learning promotes input-specific synaptic plasticity in adult-born neurons
- PMID: 25189772
- PMCID: PMC4183341
- DOI: 10.1073/pnas.1404991111
Olfactory learning promotes input-specific synaptic plasticity in adult-born neurons
Abstract
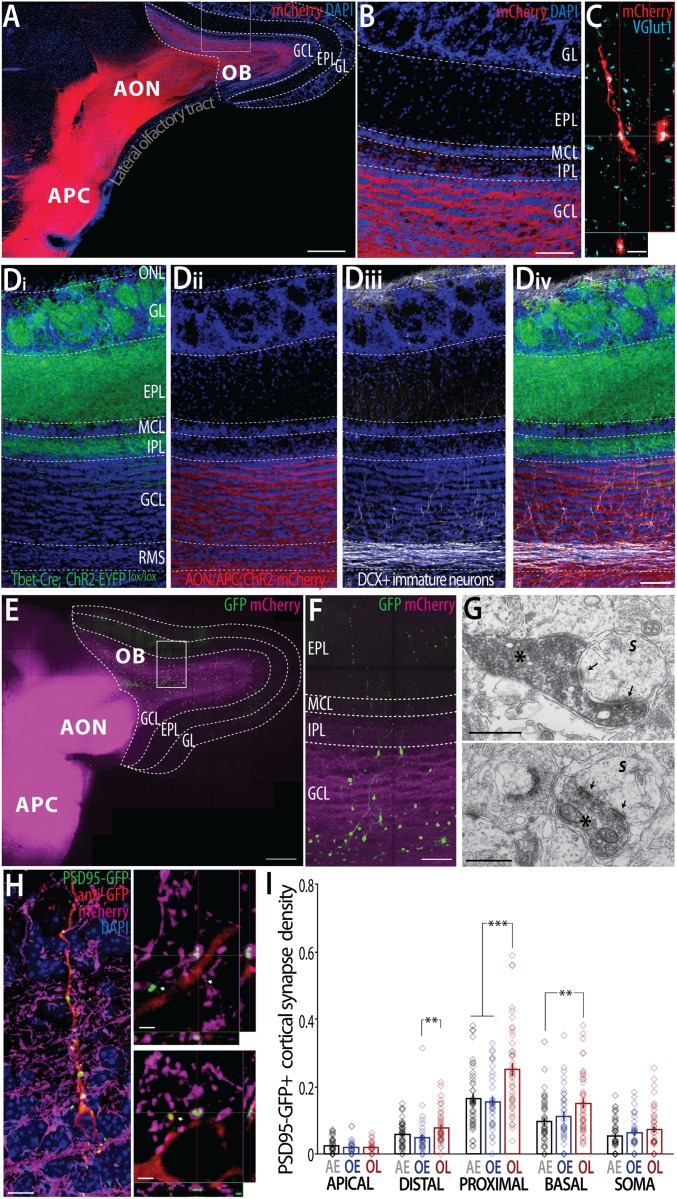
The production of new neurons in the olfactory bulb (OB) through adulthood is a major mechanism of structural and functional plasticity underlying learning-induced circuit remodeling. The recruitment of adult-born OB neurons depends not only on sensory input but also on the context in which the olfactory stimulus is received. Among the multiple steps of adult neurogenesis, the integration and survival of adult-born neurons are both strongly influenced by olfactory learning. Conversely, optogenetic stimulation of adult-born neurons has been shown to specifically improve olfactory learning and long-term memory. However, the nature of the circuit and the synaptic mechanisms underlying this reciprocal influence are not yet known. Here, we showed that olfactory learning increases the spine density in a region-restricted manner along the dendritic tree of adult-born granule cells (GCs). Anatomical and electrophysiological analysis of adult-born GCs showed that olfactory learning promotes a remodeling of both excitatory and inhibitory inputs selectively in the deep dendritic domain. Circuit mapping revealed that the malleable dendritic portion of adult-born neurons receives excitatory inputs mostly from the regions of the olfactory cortex that project back to the OB. Finally, selective optogenetic stimulation of olfactory cortical projections to the OB showed that learning strengthens these inputs onto adult-born GCs. We conclude that learning promotes input-specific synaptic plasticity in adult-born neurons, which reinforces the top-down influence from the olfactory cortex to early stages of olfactory information processing.
Keywords: cortico-bulbar projections; glutamate; inhibitory circuits; piriform cortex; sensory systems.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
