

The complexity of miRNA-mediated repression
- PMID: 25190144
- PMCID: PMC4262769
- DOI: 10.1038/cdd.2014.112
The complexity of miRNA-mediated repression
Abstract
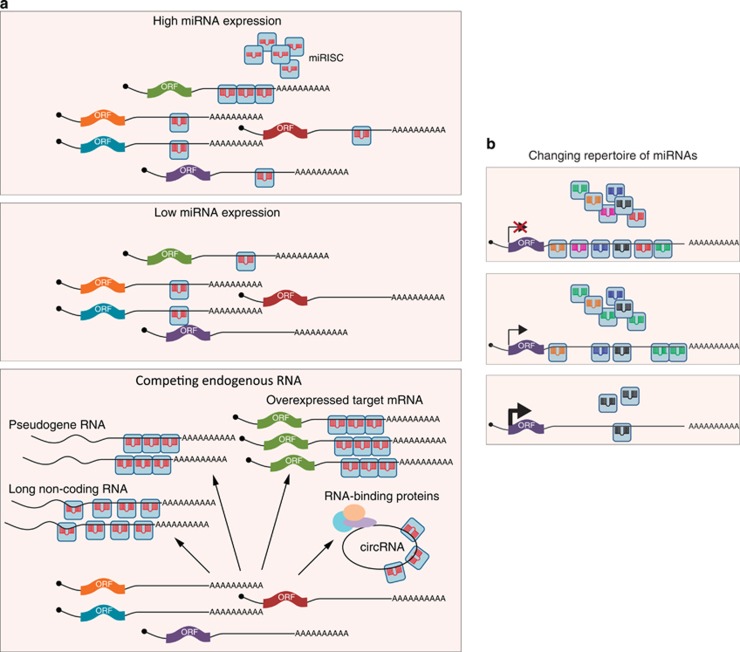
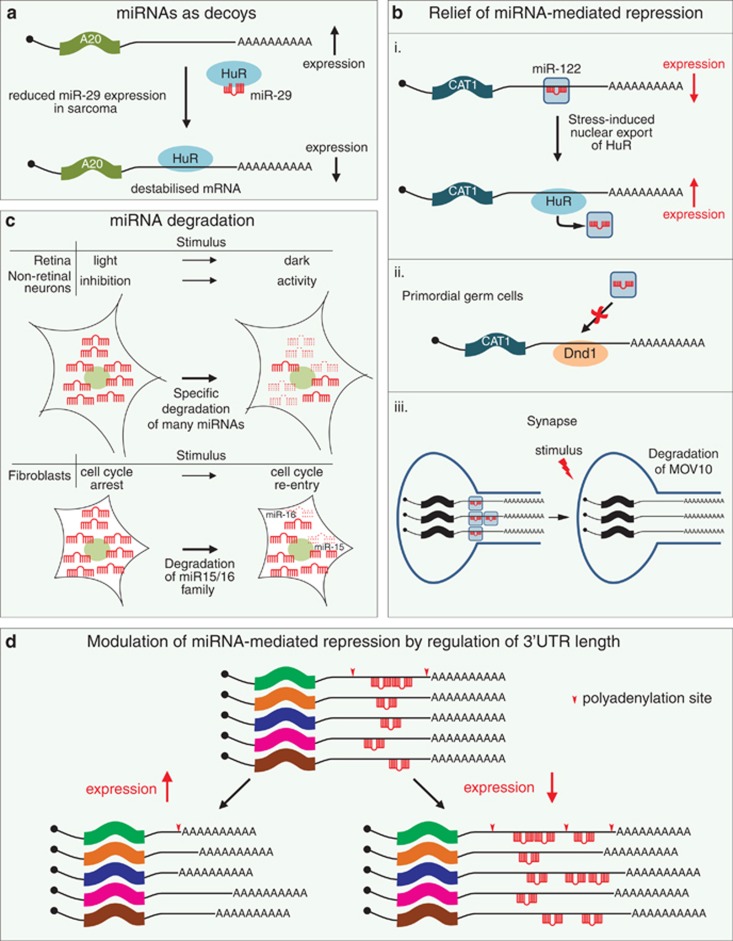
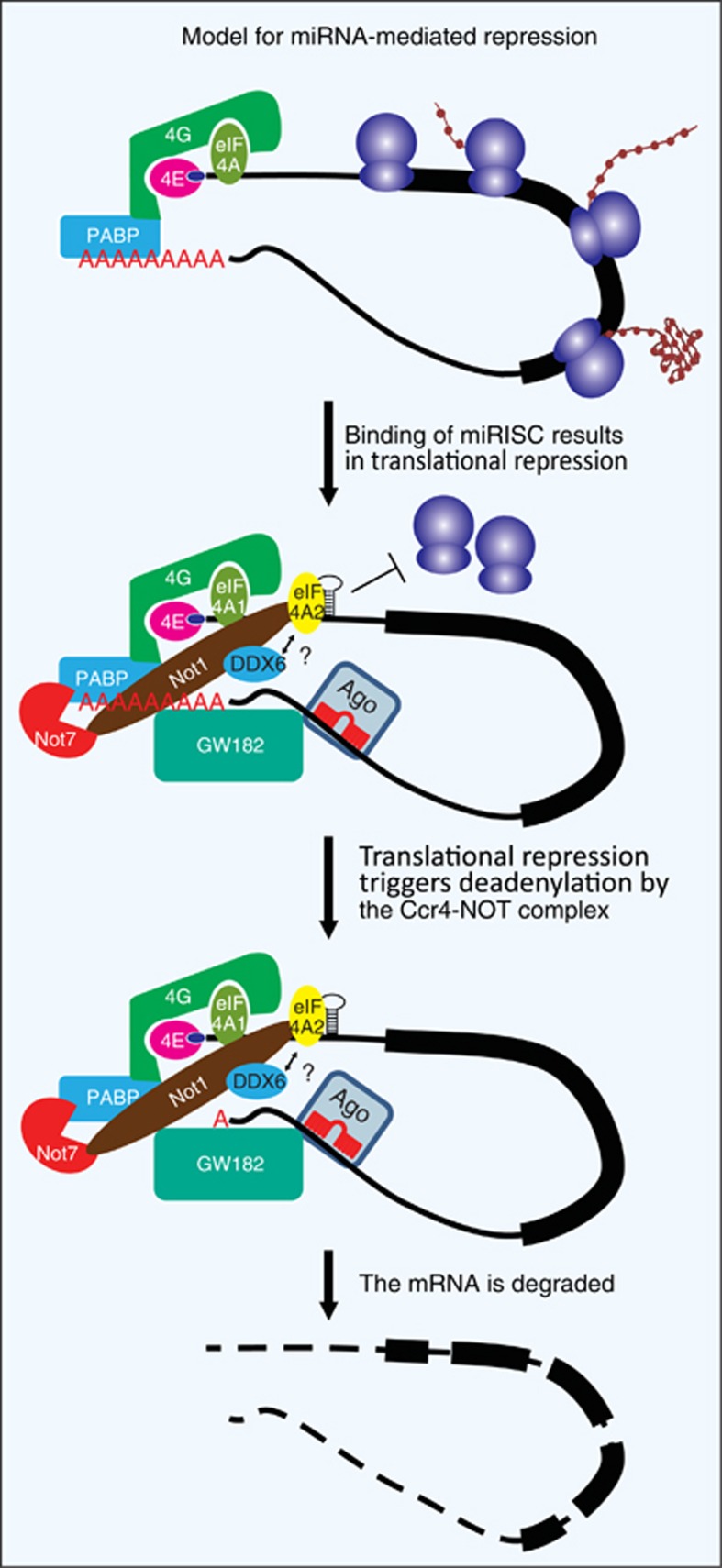
Since their discovery 20 years ago, miRNAs have attracted much attention from all areas of biology. These short (∼22 nt) non-coding RNA molecules are highly conserved in evolution and are present in nearly all eukaryotes. They have critical roles in virtually every cellular process, particularly determination of cell fate in development and regulation of the cell cycle. Although it has long been known that miRNAs bind to mRNAs to trigger translational repression and degradation, there had been much debate regarding their precise mode of action. It is now believed that translational control is the primary event, only later followed by mRNA destabilisation. This review will discuss the most recent advances in our understanding of the molecular underpinnings of miRNA-mediated repression. Moreover, we highlight the multitude of regulatory mechanisms that modulate miRNA function.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
