

Bradykinin enhances invasion of malignant glioma into the brain parenchyma by inducing cells to undergo amoeboid migration
- PMID: 25194042
- PMCID: PMC4259546
- DOI: 10.1113/jphysiol.2014.274498
Bradykinin enhances invasion of malignant glioma into the brain parenchyma by inducing cells to undergo amoeboid migration
Abstract
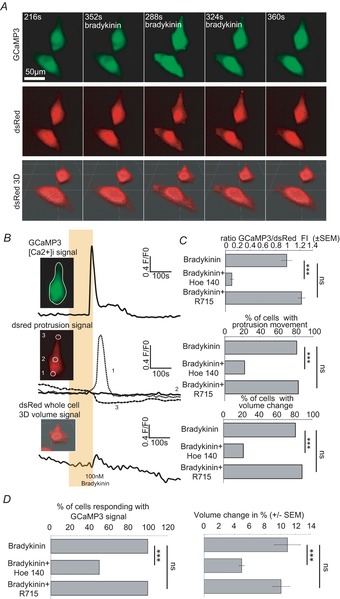
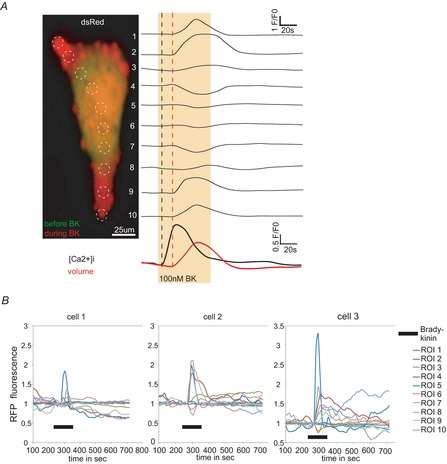
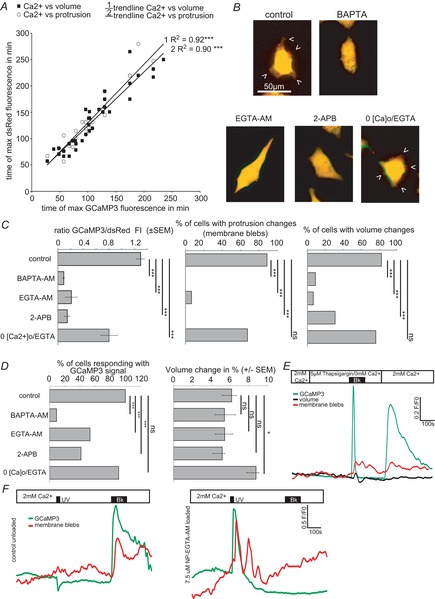
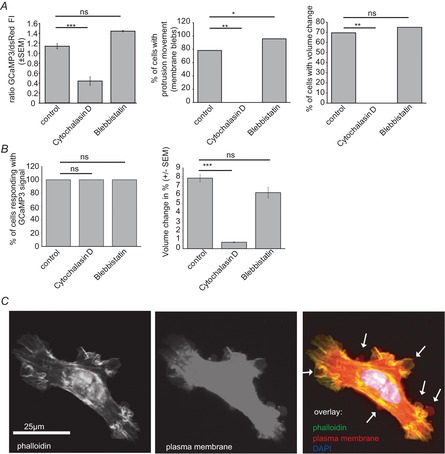
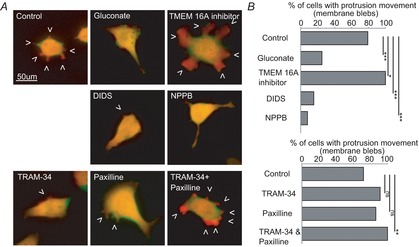
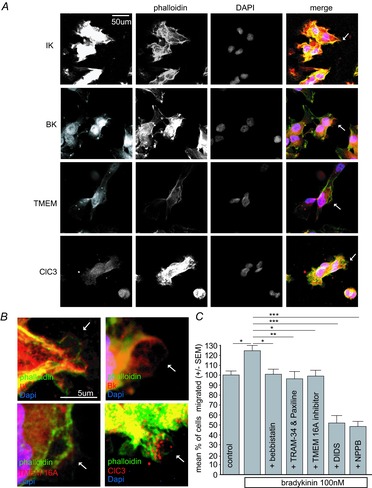
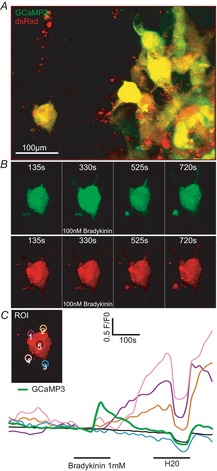
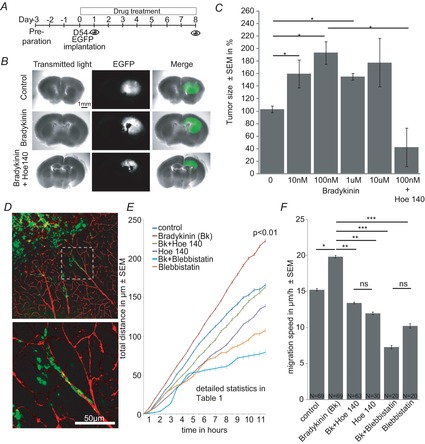
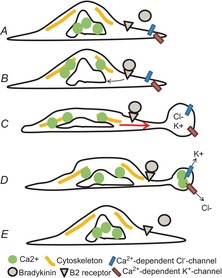
The molecular and cellular mechanisms governing cell motility and directed migration in response to the neuropeptide bradykinin are largely unknown. Here, we demonstrate that human glioma cells whose migration is guided by bradykinin generate bleb-like protrusions. We found that activation of the B2 receptor leads to a rise in free Ca(2+) from internal stores that activates actomyosin contraction and subsequent cytoplasmic flow into protrusions forming membrane blebs. Furthermore Ca(2+) activates Ca(2+)-dependent K(+) and Cl(-) channels, which participate in bleb regulation. Treatment of gliomas with bradykinin in situ increased glioma growth by increasing the speed of cell migration at the periphery of the tumour mass. To test if bleb formation is related to bradykinin-promoted glioma invasion we blocked glioma migration with blebbistatin, a blocker of myosin kinase II, which is necessary for proper bleb retraction. Our findings suggest a pivotal role of bradykinin during glioma invasion by stimulating amoeboid migration of glioma cells.
© 2014 The Authors. The Journal of Physiology © 2014 The Physiological Society.
Figures
Comment in
-
Lethal migration: the bradykinin story.J Physiol. 2014 Nov 15;592(22):4805-6. doi: 10.1113/jphysiol.2014.284455. Epub 2014 Sep 25. J Physiol. 2014. PMID: 25260635 Free PMC article. No abstract available.
References
-
- Blaser H, Reichman-Fried M, Castanon I, Dumstrei K, Marlow FL, Kawakami K, Solnica-Krezel L, Heisenberg C-P. Raz E. Migration of zebrafish primordial germ cells: a role for myosin contraction and cytoplasmic flow. Dev Cell. 2006;11:613–627. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous