

Protein dynamics during presynaptic-complex assembly on individual single-stranded DNA molecules
- PMID: 25195049
- PMCID: PMC4190069
- DOI: 10.1038/nsmb.2886
Protein dynamics during presynaptic-complex assembly on individual single-stranded DNA molecules
Abstract
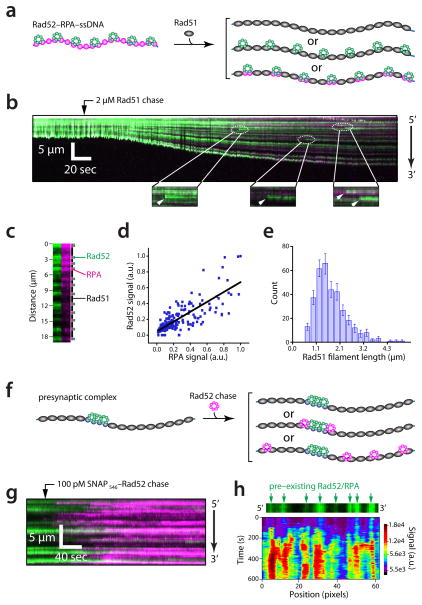
Homologous recombination is a conserved pathway for repairing double-stranded breaks, which are processed to yield single-stranded DNA overhangs that serve as platforms for presynaptic-complex assembly. Here we use single-molecule imaging to reveal the interplay between Saccharomyces cerevisiae RPA, Rad52 and Rad51 during presynaptic-complex assembly. We show that Rad52 binds RPA-ssDNA and suppresses RPA turnover, highlighting an unanticipated regulatory influence on protein dynamics. Rad51 binding extends the ssDNA, and Rad52-RPA clusters remain interspersed along the presynaptic complex. These clusters promote additional binding of RPA and Rad52. Our work illustrates the spatial and temporal progression of the association of RPA and Rad52 with the presynaptic complex and reveals a new RPA-Rad52-Rad51-ssDNA intermediate, with implications for how the activities of Rad52 and RPA are coordinated with Rad51 during the later stages of recombination.
Conflict of interest statement
The authors declare no competing financial interests.
Figures






References
-
- Mazon G, Mimitou EP, Symington LS. SnapShot: Homologous recombination in DNA double–strand break repair. Cell. 2010;142:646, 646 e1. - PubMed
-
- San Filippo J, Sung P, Klein H. Mechanism of eukaryotic homologous recombination. Annu Rev Biochem. 2008;77:229–57. - PubMed
-
- Krogh BO, Symington LS. Recombination proteins in yeast. Annu Rev Genet. 2004;38:233–71. - PubMed
-
- Cromie GA, Connelly JC, Leach DR. Recombination at double–strand breaks and DNA ends: conserved mechanisms from phage to humans. Mol Cell. 2001;8:1163–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
