

Amino acids 89-96 of Salmonella typhimurium flagellin represent the major domain responsible for TLR5-independent adjuvanticity in the humoral immune response
- PMID: 25195514
- PMCID: PMC4579647
- DOI: 10.1038/cmi.2014.76
Amino acids 89-96 of Salmonella typhimurium flagellin represent the major domain responsible for TLR5-independent adjuvanticity in the humoral immune response
Abstract
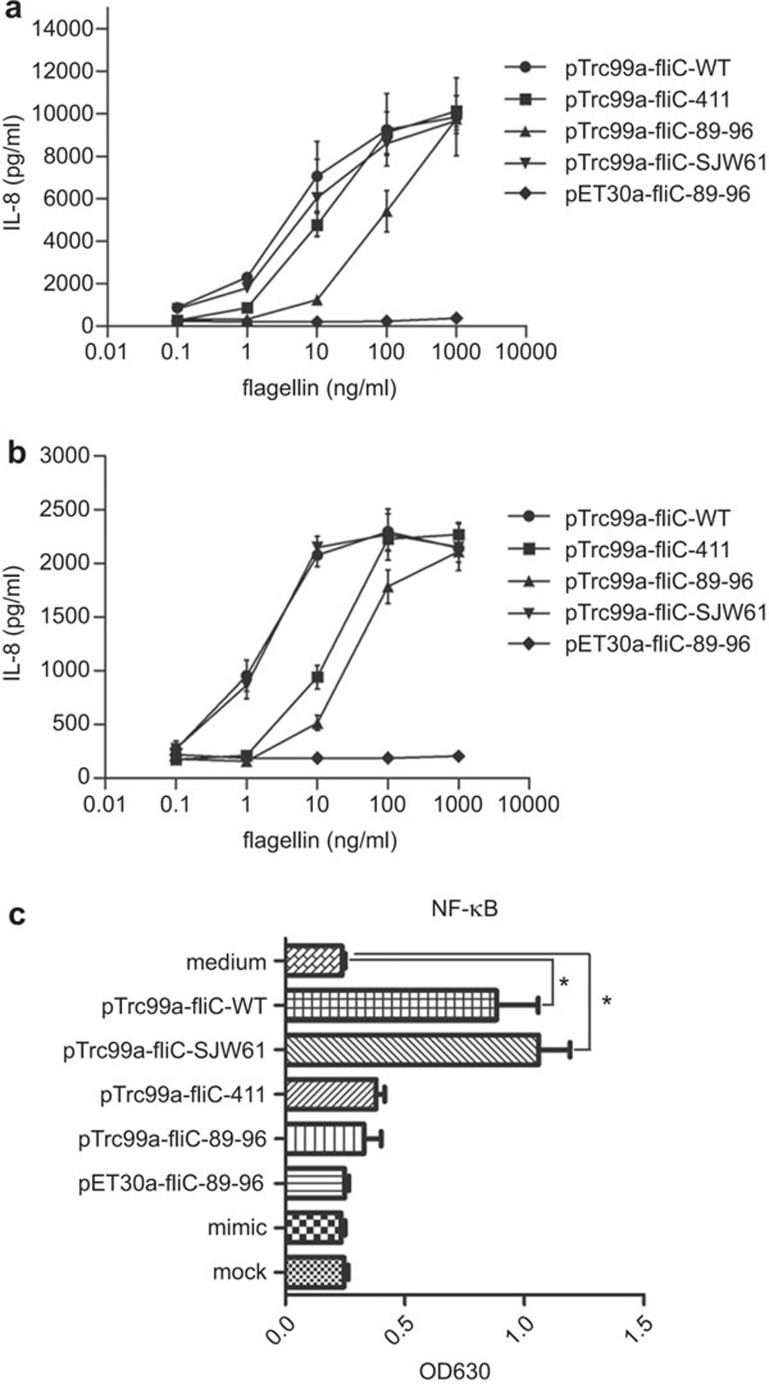
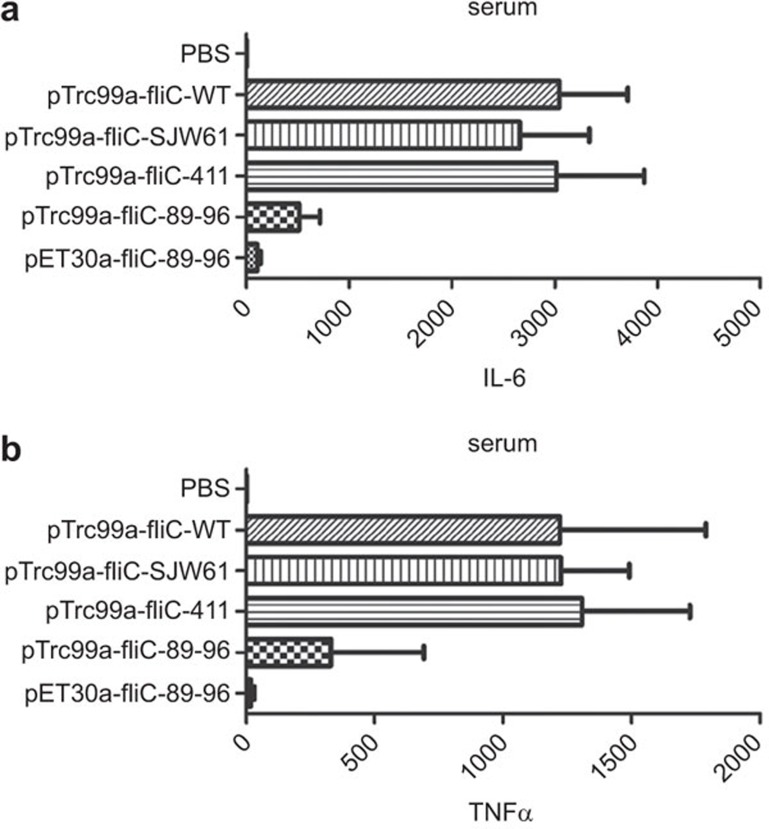
Toll-like receptor 5 (TLR5) signaling in response to flagellin is dispensable for inducing humoral immunity, but alterations of aa 89-96, the TLR5 binding site, significantly reduced the adjuvanticity of flagellin. These observations indicate that the underlying mechanism remains incompletely understood. Here, we found that the native form of Salmonella typhimurium aa 89-96-mutant flagellin extracted from flagella retains some TLR5 recognition activity, indicating that aa 89-96 is the primary, but not the only site that imparts TLR5 activity. Additionally, this mutation impaired the production of IL-1β and IL-18. Using TLR5KO mice, we found that aa 89-96 is critical for the humoral adjuvant effect, but this effect was independent of TLR5 activation triggered by this region of flagellin. In summary, our findings suggest that aa 89-96 of flagellin is not only the crucial site responsible for TLR5 recognition, but is also important for humoral immune adjuvanticity through a TLR5-independent pathway.
Figures
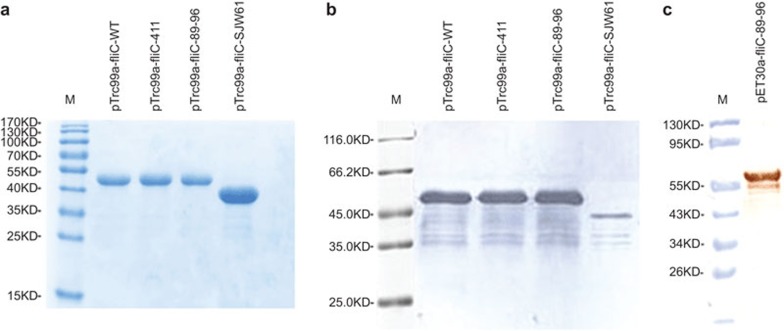



Comment in
-
Amino acids 89-96 of Salmonella flagellin: a key site for its adjuvant effect independent of the TLR5 signaling pathway.Cell Mol Immunol. 2017 Dec;14(12):1023-1025. doi: 10.1038/cmi.2017.125. Epub 2017 Nov 27. Cell Mol Immunol. 2017. PMID: 29176749 Free PMC article. No abstract available.
References
-
- 1Donnelly MA, Steiner TS. Two nonadjacent regions in enteroaggregative Escherichia coli flagellin are required for activation of Toll-like receptor 5. J Biol Chem 2002; 277: 40456–40461. - PubMed
-
- 2Eaves-Pyles TD, Wong HR, Odoms K, Pyles RB. Salmonella flagellin-dependent proinflammatory responses are localized to the conserved amino and carboxyl regions of the protein. J Immunol 2001; 167: 7009–7016. - PubMed
-
- 3Murthy KG, Deb A, Goonesekera S, Szabo C, Salzman AL. Identification of conserved domains in Salmonella muenchen flagellin that are essential for its ability to activate TLR5 and to induce an inflammatory response in vitro. J Biol Chem 2004; 279: 5667–5675. - PubMed
-
- 5Miao EA, Andersen-Nissen E, Warren SE, Aderem A. TLR5 and Ipaf: dual sensors of bacterial flagellin in the innate immune system. Semin Immunopathol 2007; 29: 275–288. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
