

Latent and active abPPO4 mushroom tyrosinase cocrystallized with hexatungstotellurate(VI) in a single crystal
- PMID: 25195745
- PMCID: PMC4157443
- DOI: 10.1107/S1399004714013777
Latent and active abPPO4 mushroom tyrosinase cocrystallized with hexatungstotellurate(VI) in a single crystal
Abstract
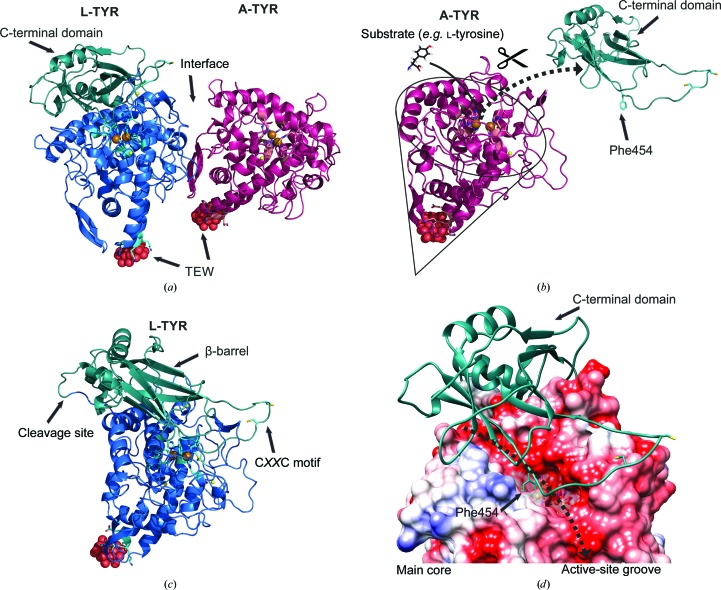
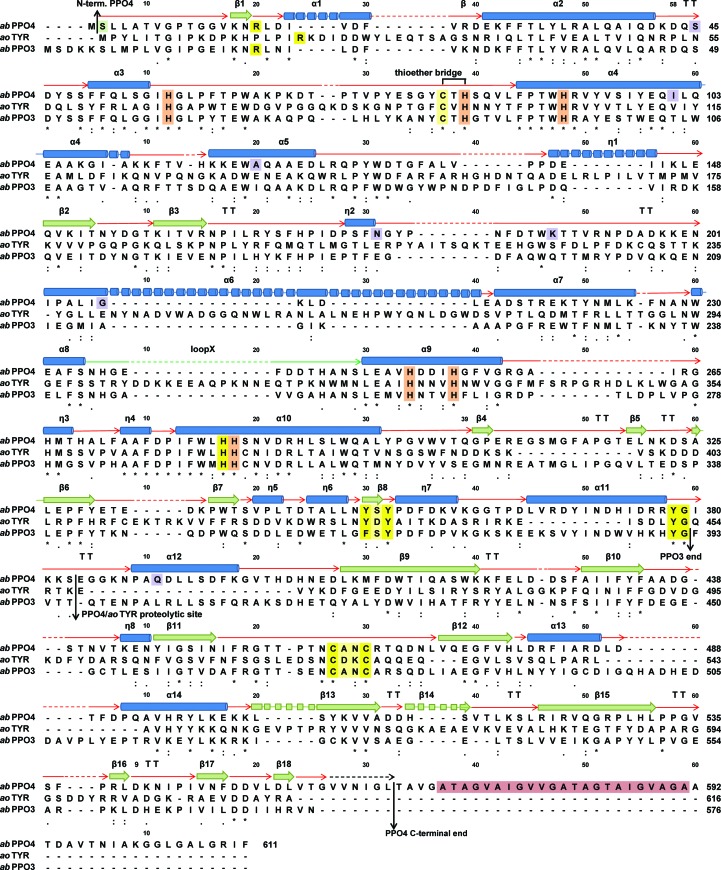
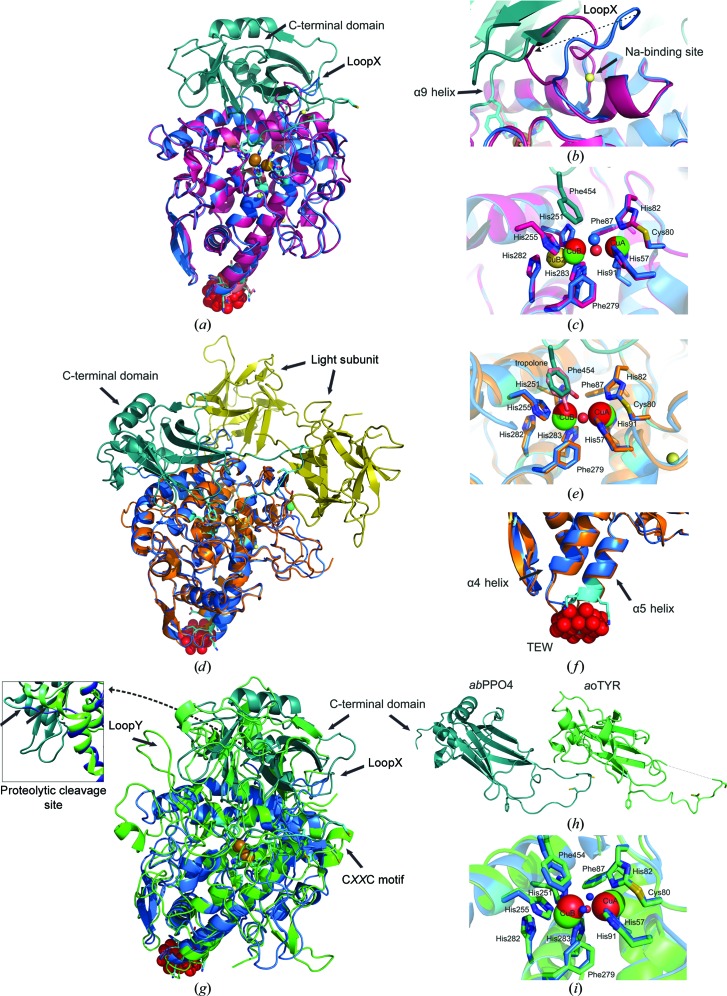
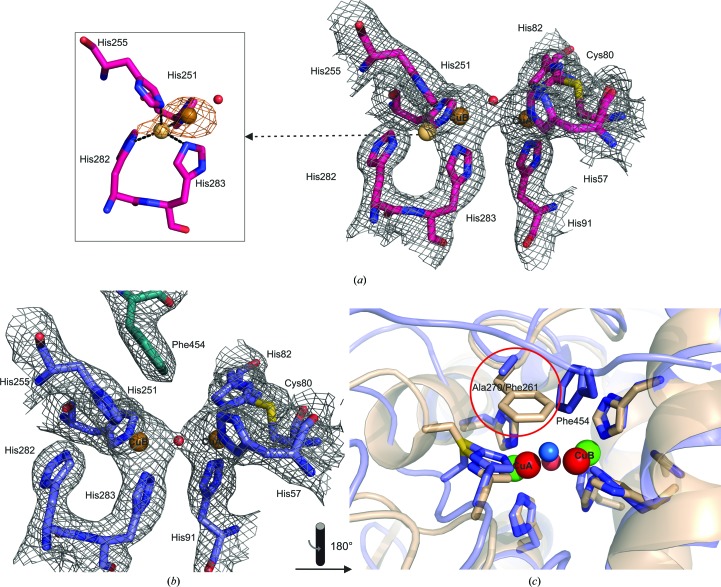
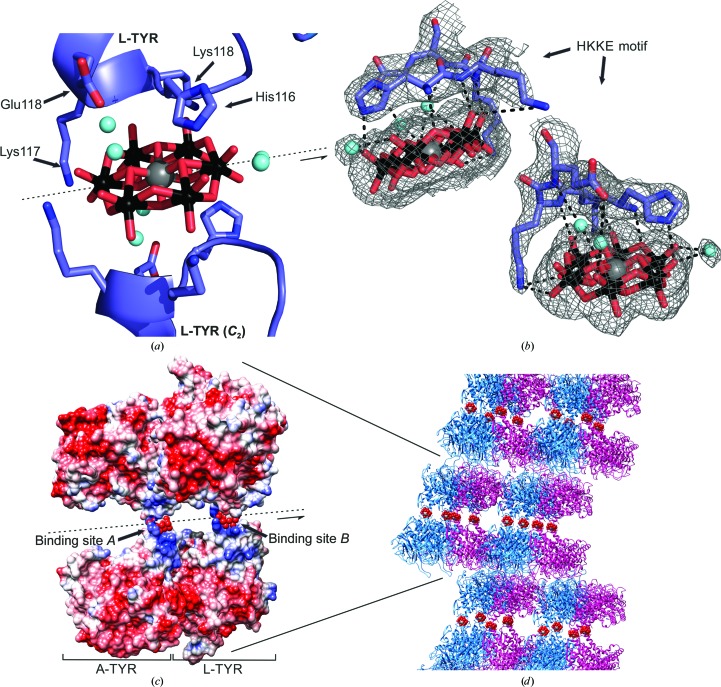
Tyrosinases, bifunctional metalloenzymes, catalyze the oxidation of monophenols and o-diphenols to o-quinones, the precursor compounds of the brown-coloured pigment melanin. In eukaryotic organisms, tyrosinases are expressed as latent zymogens that have to be proteolytically cleaved in order to form highly active enzymes. This activation mechanism, known as the tyrosinase maturation process, has scientific and industrial significance with respect to biochemical and technical applications of the enzyme. Here, not only the first crystal structure of the mushroom tyrosinase abPPO4 is presented in its active form (Ser2-Ser383) and in its 21 kDa heavier latent form (Ser2-Thr545), but furthermore the simultaneous presence of both forms within one single-crystal structure is shown. This allows for a simple approach to investigate the transition between these two forms. Isoform abPPO4 was isolated and extensively purified from the natural source (Agaricus bisporus), which contains a total of six polyphenol oxidases (PPOs). The enzyme formed crystals (diffracting to a resolution of 2.76 Å) owing to the employment of the 6-tungstotellurate(VI) salt (Na6[TeW6O24]·22H2O) as a cocrystallization agent. Two of these disc-shaped Anderson-type polyoxoanions [TeW6O24](6-) separate two asymmetric units comprising one crystallographic heterodimer of abPPO4, thus resulting in very interesting crystal packing.
Keywords: Agaricus bisporus; abPPO4; polyphenol oxidase; tyrosinase.
Figures

References
-
- Adams, P. D. et al. (2010). Acta Cryst. D66, 213–221.
-
- Bourquelot, E. & Bertrand, G. (1895). C. R. Soc. Biol. 47, 582–584.
-
- Corey, R. B., Stanford, R. H., Marsh, R. E., Leung, Y. C. & Kay, L. M. (1962). Acta Cryst. 15, 1157–1163.
-
- Cuff, M. E., Miller, K. I., van Holde, K. E. & Hendrickson, W. A. (1998). J. Mol. Biol. 278, 855–870. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
