

Blood-gene expression reveals reduced circadian rhythmicity in individuals resistant to sleep deprivation
- PMID: 25197809
- PMCID: PMC4173916
- DOI: 10.5665/sleep.4064
Blood-gene expression reveals reduced circadian rhythmicity in individuals resistant to sleep deprivation
Abstract
Study objectives: To address whether changes in gene expression in blood cells with sleep loss are different in individuals resistant and sensitive to sleep deprivation.
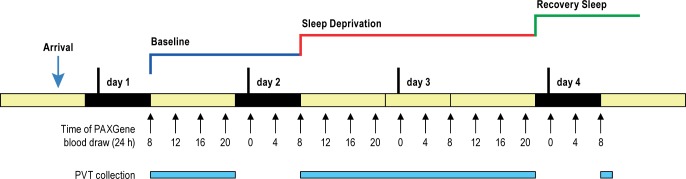
Design: Blood draws every 4 h during a 3-day study: 24-h normal baseline, 38 h of continuous wakefulness and subsequent recovery sleep, for a total of 19 time-points per subject, with every 2-h psychomotor vigilance task (PVT) assessment when awake.
Setting: Sleep laboratory.
Participants: Fourteen subjects who were previously identified as behaviorally resistant (n = 7) or sensitive (n = 7) to sleep deprivation by PVT.
Intervention: Thirty-eight hours of continuous wakefulness.
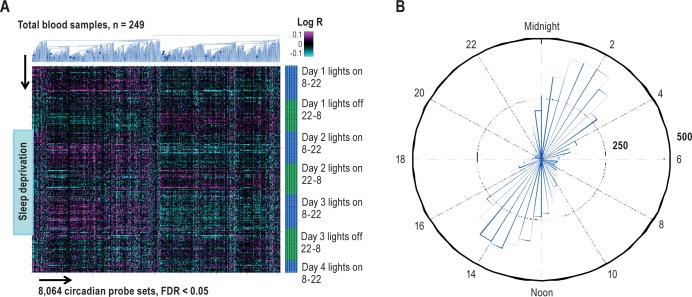
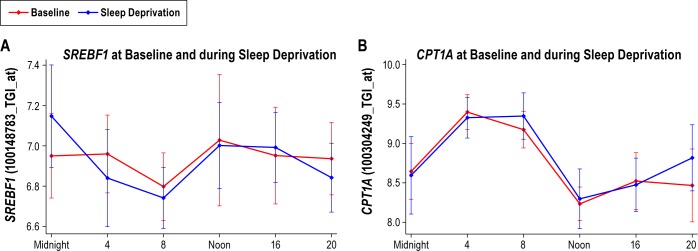
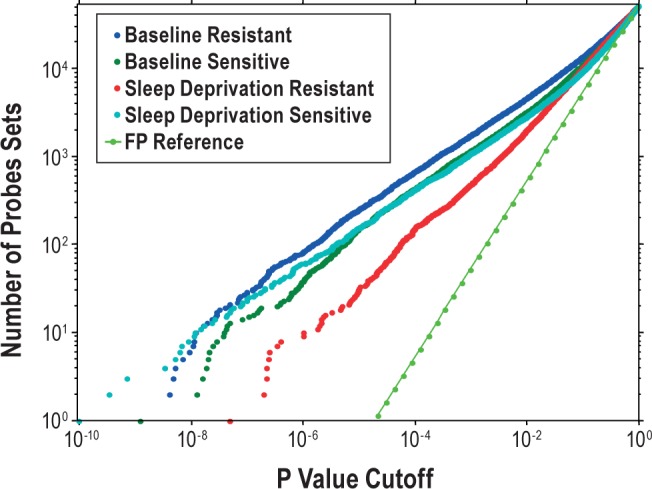
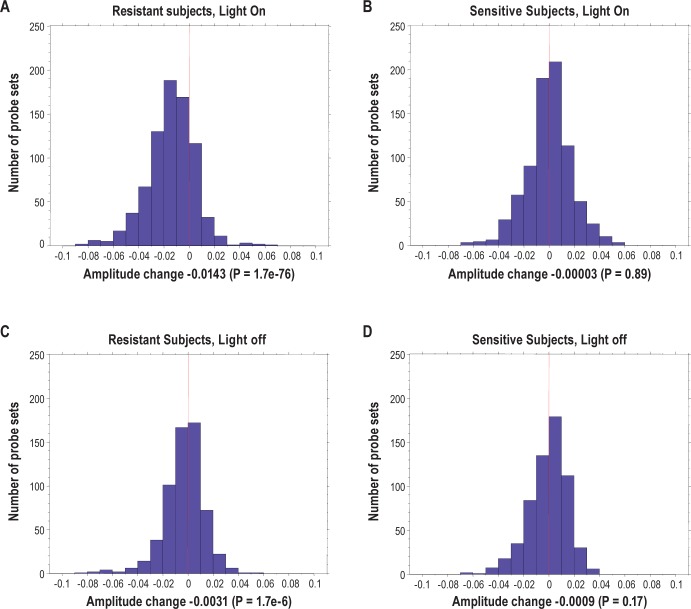
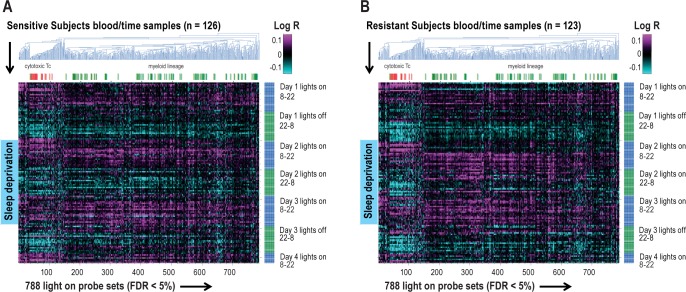
Measurements and results: We found 4,481 unique genes with a significant 24-h diurnal rhythm during a normal sleep-wake cycle in blood (false discovery rate [FDR] < 5%). Biological pathways were enriched for biosynthetic processes during sleep. After accounting for circadian effects, two genes (SREBF1 and CPT1A, both involved in lipid metabolism) exhibited small, but significant, linear changes in expression with the duration of sleep deprivation (FDR < 5%). The main change with sleep deprivation was a reduction in the amplitude of the diurnal rhythm of expression of normally cycling probe sets. This reduction was noticeably higher in behaviorally resistant subjects than sensitive subjects, at any given P value. Furthermore, blood cell type enrichment analysis showed that the expression pattern difference between sensitive and resistant subjects is mainly found in cells of myeloid origin, such as monocytes.
Conclusion: Individual differences in behavioral effects of sleep deprivation are associated with differences in diurnal amplitude of gene expression for genes that show circadian rhythmicity.
Keywords: circadian rhythm; gene expression; microarray analysis; psychomotor vigilance test; sleep deprivation.
© 2014 Associated Professional Sleep Societies, LLC.
Figures
Comment in
-
Resisting sleep deprivation by breaking the link between sleep and circadian rhythms.Sleep. 2014 Oct 1;37(10):1581-2. doi: 10.5665/sleep.4056. Sleep. 2014. PMID: 25197802 Free PMC article. No abstract available.
References
-
- Leger D, Roscoat E, Bayon V, Guignard R, Paquereau J, Beck F. Short sleep in young adults: Insomnia or sleep debt? Prevalence and clinical description of short sleep in a representative sample of 1004 young adults from France. Sleep Med. 2011;12:454–62. - PubMed
-
- Van Dongen HP, Baynard MD, Maislin G, Dinges DF. Systematic interindividual differences in neurobehavioral impairment from sleep loss: evidence of trait-like differential vulnerability. Sleep. 2004;27:423–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
