

Chromosomal translocations in human cells are generated by canonical nonhomologous end-joining
- PMID: 25201414
- PMCID: PMC4398060
- DOI: 10.1016/j.molcel.2014.08.002
Chromosomal translocations in human cells are generated by canonical nonhomologous end-joining
Abstract
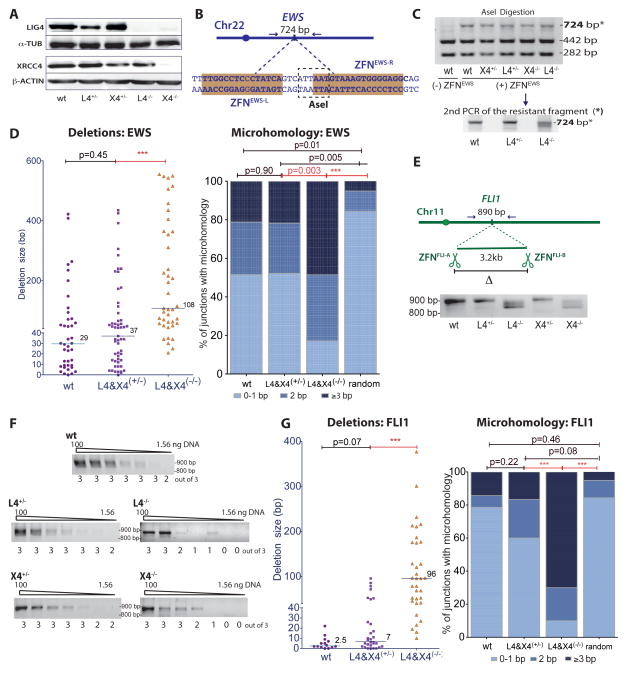
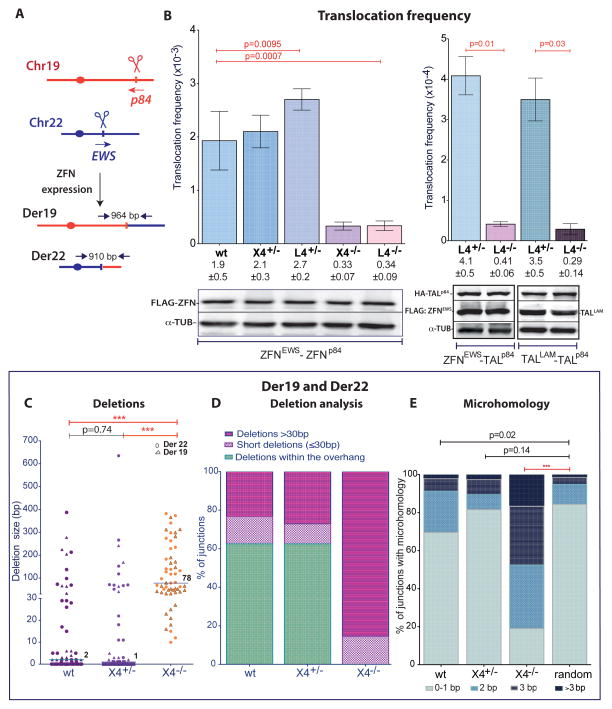
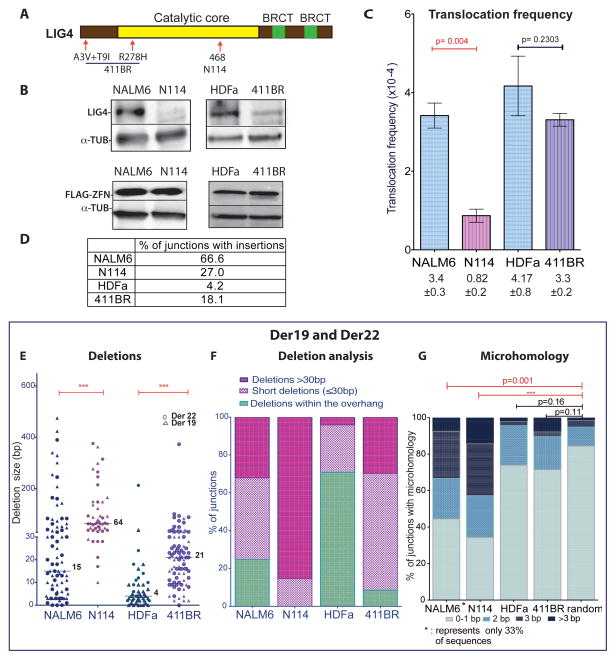
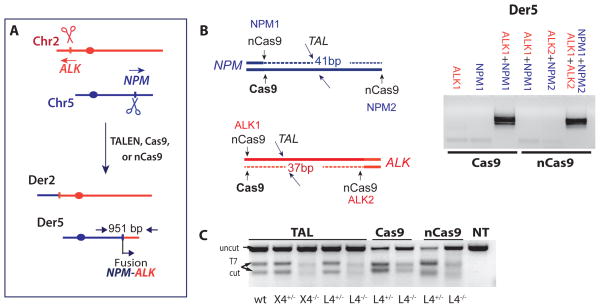
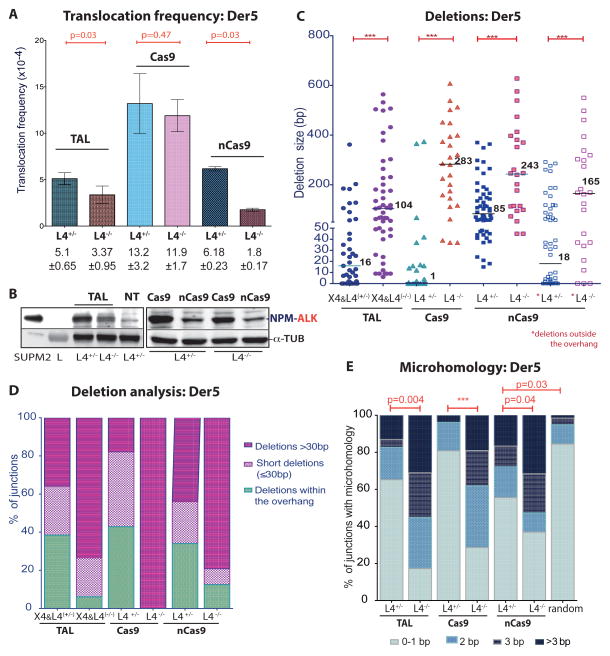
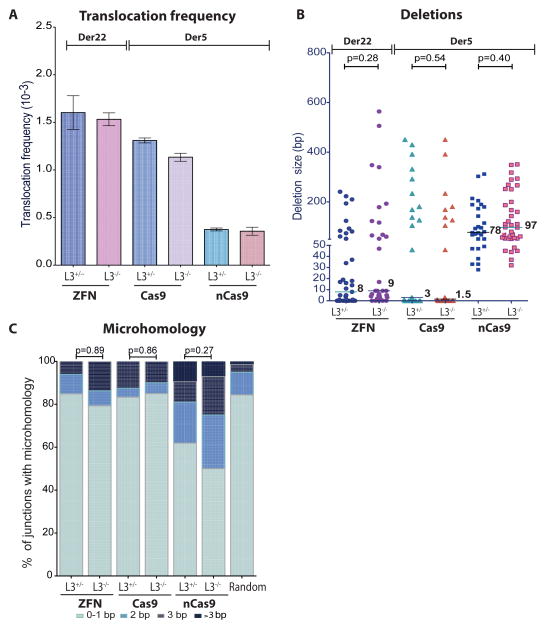
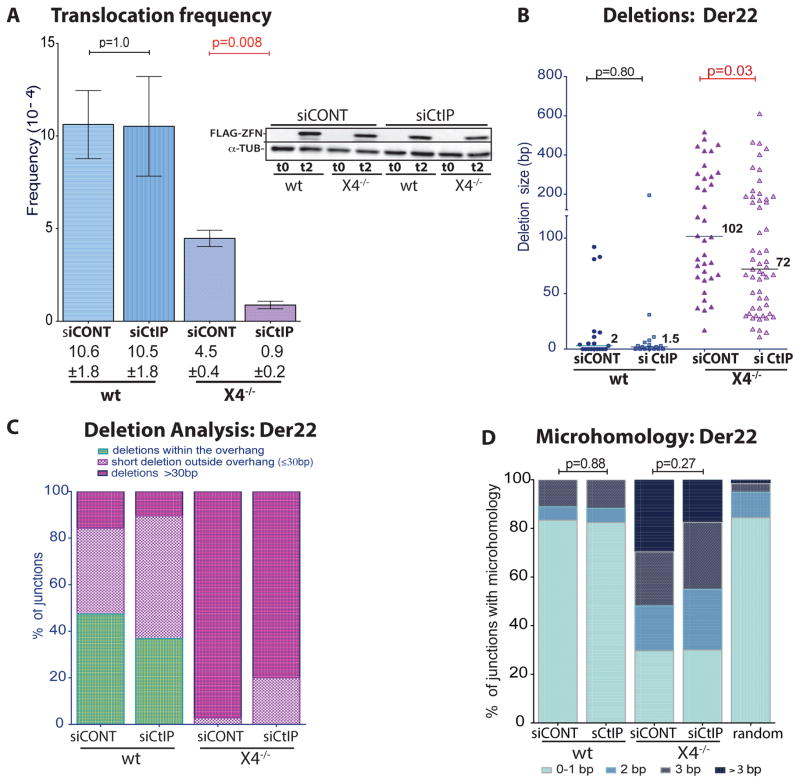
Breakpoint junctions of the chromosomal translocations that occur in human cancers display hallmarks of nonhomologous end-joining (NHEJ). In mouse cells, translocations are suppressed by canonical NHEJ (c-NHEJ) components, which include DNA ligase IV (LIG4), and instead arise from alternative NHEJ (alt-NHEJ). Here we used designer nucleases (ZFNs, TALENs, and CRISPR/Cas9) to introduce DSBs on two chromosomes to study translocation joining mechanisms in human cells. Remarkably, translocations were altered in cells deficient for LIG4 or its interacting protein XRCC4. Translocation junctions had significantly longer deletions and more microhomology, indicative of alt-NHEJ. Thus, unlike mouse cells, translocations in human cells are generated by c-NHEJ. Human cancer translocations induced by paired Cas9 nicks also showed a dependence on c-NHEJ, despite having distinct joining characteristics. These results demonstrate an unexpected and striking species-specific difference for common genomic rearrangements associated with tumorigenesis.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Boboila C, Alt FW, Schwer B. Classical and alternative end-joining pathways for repair of lymphocyte-specific and general DNA double-strand breaks. Adv Immunol. 2012a;116:1–49. - PubMed
-
- Boboila C, Jankovic M, Yan CT, Wang JH, Wesemann DR, Zhang T, Fazeli A, Feldman L, Nussenzweig A, Nussenzweig M, et al. Alternative end-joining catalyzes robust IgH locus deletions and translocations in the combined absence of ligase 4 and Ku70. Proc Natl Acad Sci USA. 2010;107:3034–3039. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
