

Current perspectives on the hormonal control of seed development in Arabidopsis and maize: a focus on auxin
- PMID: 25202316
- PMCID: PMC4142864
- DOI: 10.3389/fpls.2014.00412
Current perspectives on the hormonal control of seed development in Arabidopsis and maize: a focus on auxin
Abstract
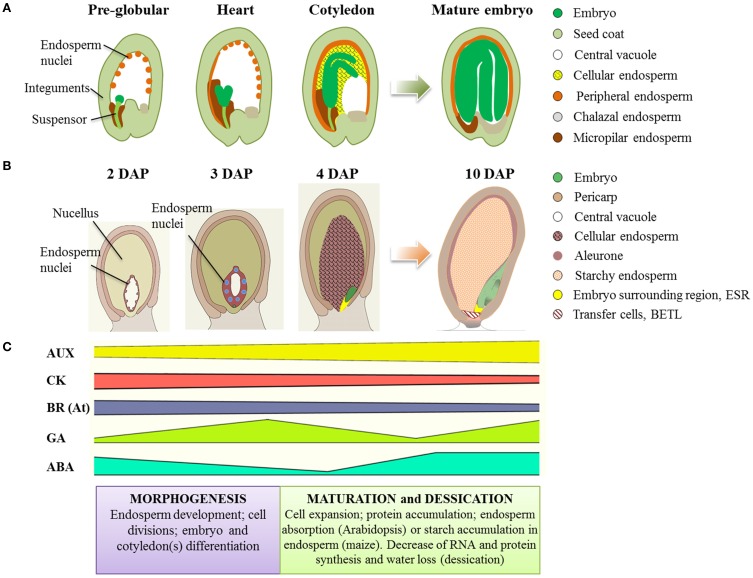
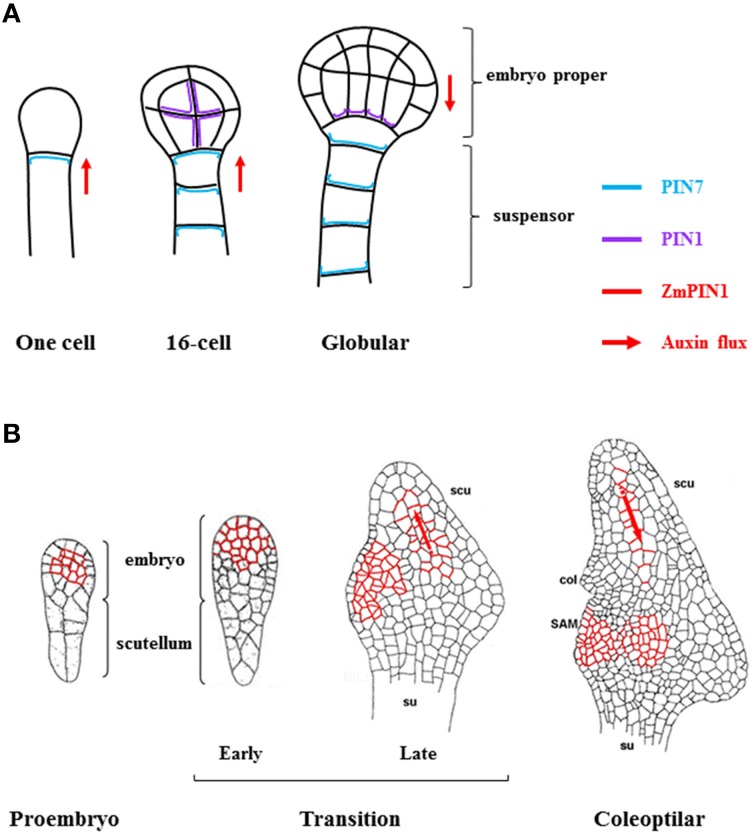
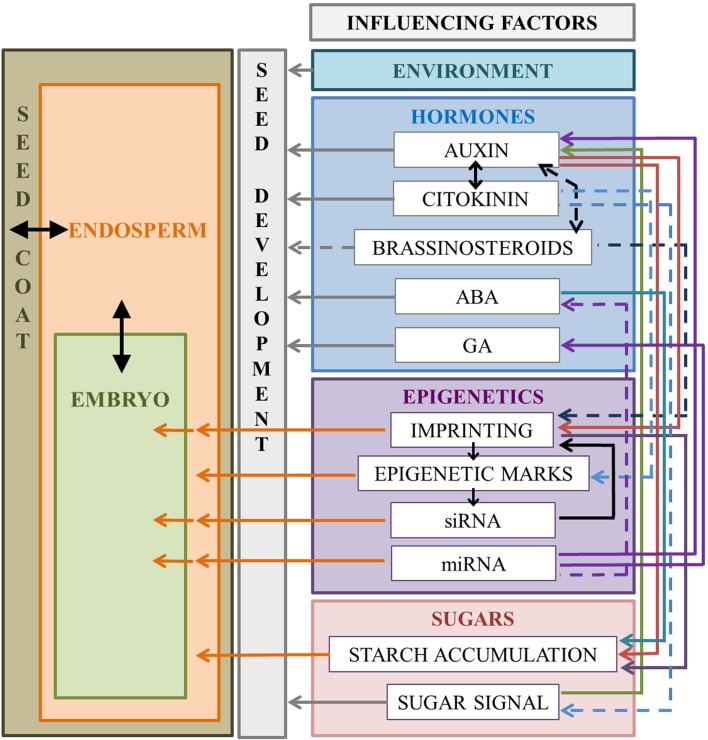
The seed represents the unit of reproduction of flowering plants, capable of developing into another plant, and to ensure the survival of the species under unfavorable environmental conditions. It is composed of three compartments: seed coat, endosperm and embryo. Proper seed development depends on the coordination of the processes that lead to seed compartments differentiation, development and maturation. The coordination of these processes is based on the constant transmission/perception of signals by the three compartments. Phytohormones constitute one of these signals; gradients of hormones are generated in the different seed compartments, and their ratios comprise the signals that induce/inhibit particular processes in seed development. Among the hormones, auxin seems to exert a central role, as it is the only one in maintaining high levels of accumulation from fertilization to seed maturation. The gradient of auxin generated by its PIN carriers affects several processes of seed development, including pattern formation, cell division and expansion. Despite the high degree of conservation in the regulatory mechanisms that lead to seed development within the Spermatophytes, remarkable differences exist during seed maturation between Monocots and Eudicots species. For instance, in Monocots the endosperm persists until maturation, and constitutes an important compartment for nutrients storage, while in Eudicots it is reduced to a single cell layer, as the expanding embryo gradually replaces it during the maturation. This review provides an overview of the current knowledge on hormonal control of seed development, by considering the data available in two model plants: Arabidopsis thaliana, for Eudicots and Zea mays L., for Monocots. We will emphasize the control exerted by auxin on the correct progress of seed development comparing, when possible, the two species.
Keywords: Arabidopsis; auxin; embryo; endosperm; maize; phytohormones; seed development.
Figures
Similar articles
-
Hormonal responses during early embryogenesis in maize.Biochem Soc Trans. 2014 Apr;42(2):325-31. doi: 10.1042/BST20130260. Biochem Soc Trans. 2014. PMID: 24646239
-
Auxin production in the endosperm drives seed coat development in Arabidopsis.Elife. 2016 Nov 16;5:e20542. doi: 10.7554/eLife.20542. Elife. 2016. PMID: 27848912 Free PMC article.
-
The role of PIN auxin efflux carriers in polar auxin transport and accumulation and their effect on shaping maize development.Mol Plant. 2012 Jul;5(4):787-98. doi: 10.1093/mp/ssr103. Epub 2011 Dec 19. Mol Plant. 2012. PMID: 22186966 Review.
-
ZmPIN1-mediated auxin transport is related to cellular differentiation during maize embryogenesis and endosperm development.Plant Physiol. 2010 Mar;152(3):1373-90. doi: 10.1104/pp.109.150193. Epub 2009 Dec 31. Plant Physiol. 2010. PMID: 20044449 Free PMC article.
-
Genetic analysis as a tool to investigate the molecular mechanisms underlying seed development in maize.Ann Bot. 2005 Sep;96(3):353-62. doi: 10.1093/aob/mci187. Epub 2005 Jul 5. Ann Bot. 2005. PMID: 15998629 Free PMC article. Review.
Cited by
-
Expression patterns of Brassica napus genes implicate IPT, CKX, sucrose transporter, cell wall invertase, and amino acid permease gene family members in leaf, flower, silique, and seed development.J Exp Bot. 2015 Aug;66(16):5067-82. doi: 10.1093/jxb/erv133. Epub 2015 Apr 4. J Exp Bot. 2015. PMID: 25873685 Free PMC article.
-
Genome Wide Analysis of the Transcriptional Profiles in Different Regions of the Developing Rice Grains.Rice (N Y). 2020 Sep 7;13(1):62. doi: 10.1186/s12284-020-00421-4. Rice (N Y). 2020. PMID: 32894395 Free PMC article.
-
Systems biology of seeds: decoding the secret of biochemical seed factories for nutritional security.3 Biotech. 2018 Nov;8(11):460. doi: 10.1007/s13205-018-1483-9. Epub 2018 Oct 24. 3 Biotech. 2018. PMID: 30370201 Free PMC article. Review.
-
Cytidinediphosphate diacylglycerol synthase-Mediated phosphatidic acid metabolism is crucial for early embryonic development of Arabidopsis.PLoS Genet. 2022 Jul 25;18(7):e1010320. doi: 10.1371/journal.pgen.1010320. eCollection 2022 Jul. PLoS Genet. 2022. PMID: 35877676 Free PMC article.
-
Into the Seed: Auxin Controls Seed Development and Grain Yield.Int J Mol Sci. 2020 Feb 28;21(5):1662. doi: 10.3390/ijms21051662. Int J Mol Sci. 2020. PMID: 32121296 Free PMC article. Review.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources