

Epithelial homeostasis
- PMID: 25202877
- PMCID: PMC4196707
- DOI: 10.1016/j.cub.2014.06.068
Epithelial homeostasis
Abstract
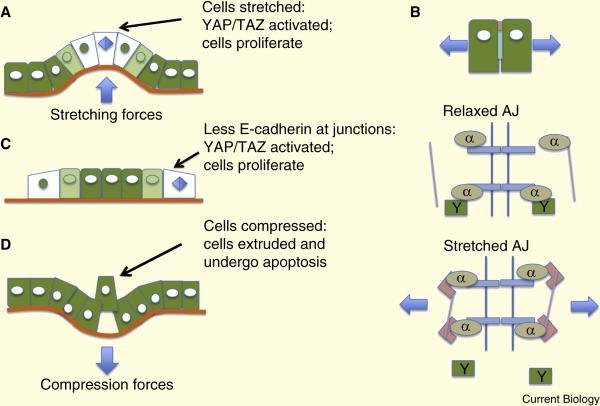
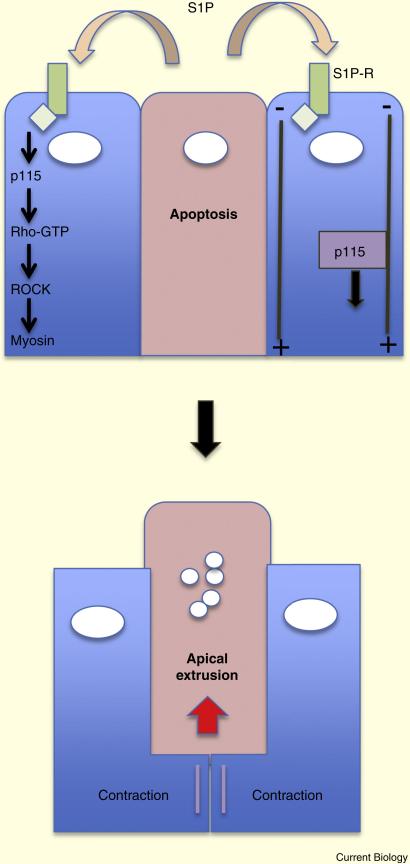
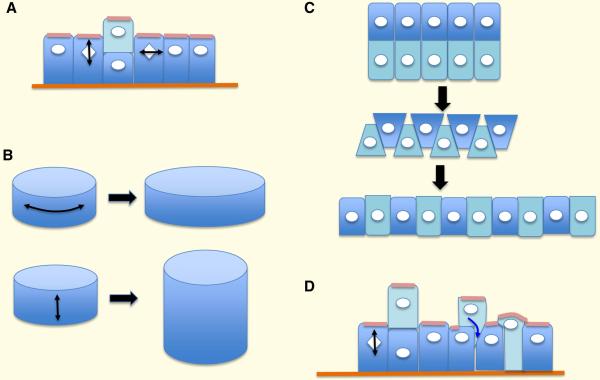
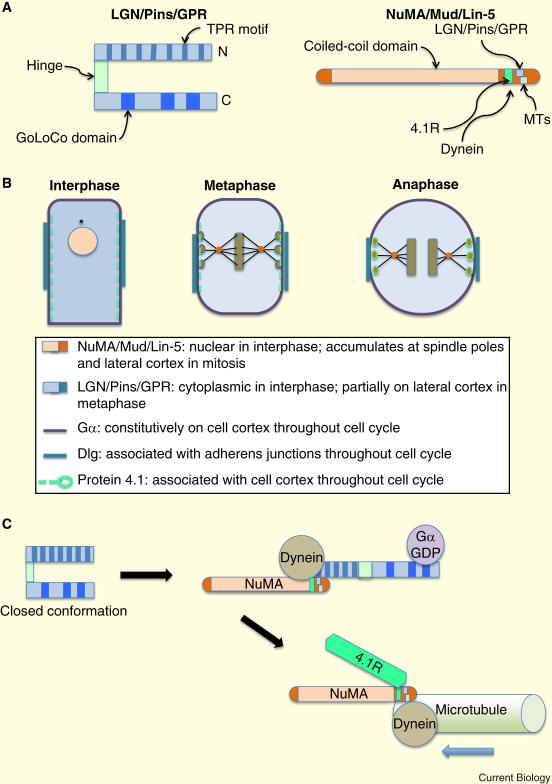
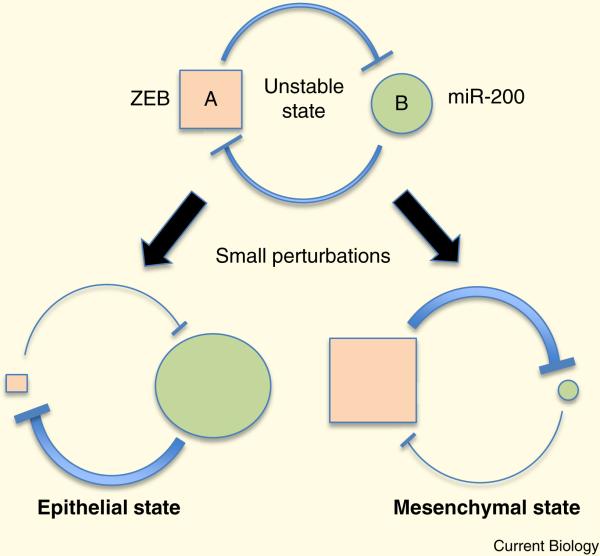
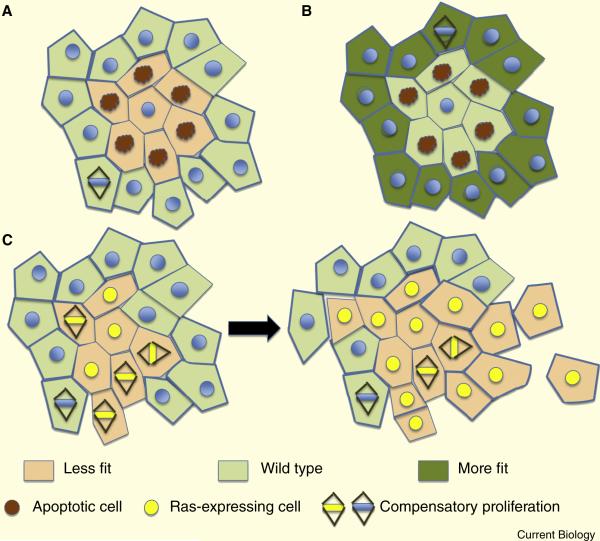
Epithelia form intelligent, dynamic barriers between the external environment and an organism's interior. Intercellular cadherin-based adhesions adapt and respond to mechanical forces and cell density, while tight junctions flexibly control diffusion both within the plasma membrane and between adjacent cells. Epithelial integrity and homeostasis are of central importance to survival, and mechanisms have evolved to ensure these processes are maintained during growth and in response to damage. For instance, cell competition surveys the fitness of cells within epithelia and removes the less fit; extrusion or delamination can remove apoptotic or defective cells from the epithelial sheet and can restore homeostasis when an epithelial layer becomes too crowded; spindle orientation ensures two-dimensional growth in simple epithelia and controls stratification in complex epithelia; and transition to a mesenchymal phenotype enables active escape from an epithelial layer. This review will discuss these various mechanisms and consider how they are subverted in disease.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Sato T, Clevers H. Growing self-organizing mini-guts from a single intestinal stem cell: mechanism and applications. Science. 2013;340:1190–1194. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
