

Construction of a chimeric biosynthetic pathway for the de novo biosynthesis of rosmarinic acid in Escherichia coli
- PMID: 25205019
- PMCID: PMC4264569
- DOI: 10.1002/cbic.201402275
Construction of a chimeric biosynthetic pathway for the de novo biosynthesis of rosmarinic acid in Escherichia coli
Abstract
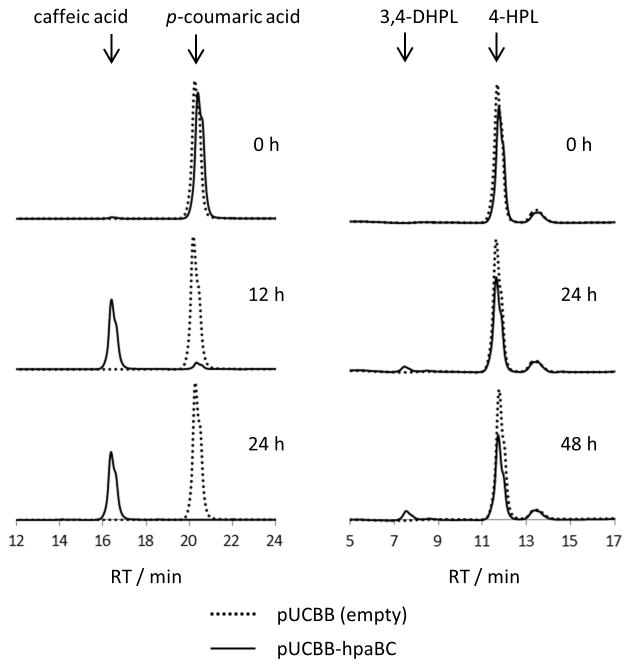
Hydroxycinnamic acid esters (HCEs) are widely-distributed phenylpropanoid-derived plant natural products. Rosmarinic acid (RA), the most well-known HCE, shows promise as a treatment for cancer and neurological disorders. In contrast to extraction from plant material or plant cell culture, microbial production of HCEs could be a sustainable, controlled means of production. Through the overexpression of a six-enzyme chimeric bacterial and plant pathway, we show the de novo biosynthesis of RA, and the related HCE isorinic acid (IA), in Escherichia coli. Probing the pathway through precursor supplementation showed several potential pathway bottlenecks. We demonstrated HCE biosynthesis using three plant rosmarinic acid synthase (RAS) orthologues, which exhibited different levels of HCE biosynthesis but produced the same ratio of IA to RA. This work serves as a proof-of-concept for a microbial production platform for HCEs by using a modular biosynthetic approach to access diverse natural and non-natural HCEs.
Keywords: biosynthesis; hydroxycinnamic acid esters; metabolic engineering; natural products; rosmarinic acid.
© 2014 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Figures






References
-
- Petersen M, Simmonds MS. Phytochemistry. 2003;62:121–125. - PubMed
-
- Bulgakov VP, Inyushkina YV, Fedoreyev SA. Crit Rev Biotechnol. 2012;32:203–217. - PubMed
-
- Moon DO, Kim MO, Lee JD, Choi YH, Kim GY. Cancer Lett. 2010;288:183–191. - PubMed
-
- Paluszczak J, Krajka-Kuzniak V, Baer-Dubowska W. Toxicol Lett. 2010;192:119–125. - PubMed
-
- Xu Y, Jiang Z, Ji G, Liu J. Planta Med. 2010;76:956–962. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
