

Ball python nidovirus: a candidate etiologic agent for severe respiratory disease in Python regius
- PMID: 25205093
- PMCID: PMC4173777
- DOI: 10.1128/mBio.01484-14
Ball python nidovirus: a candidate etiologic agent for severe respiratory disease in Python regius
Abstract
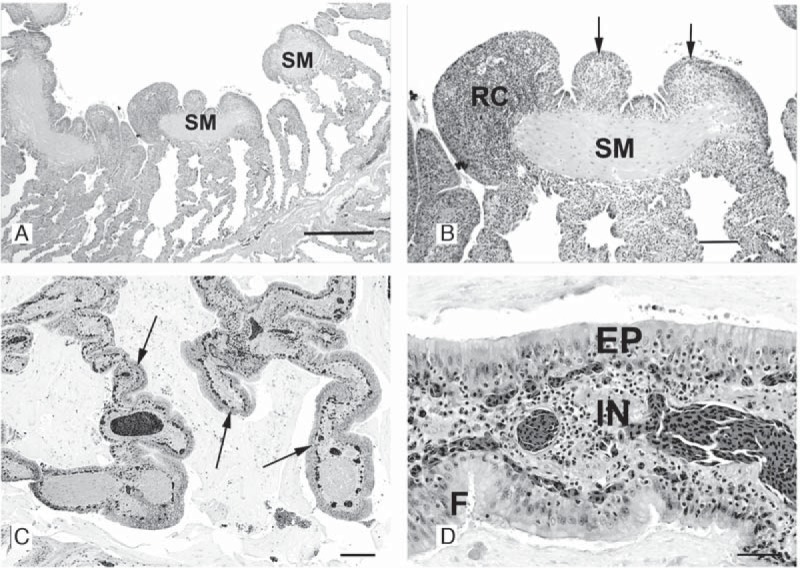
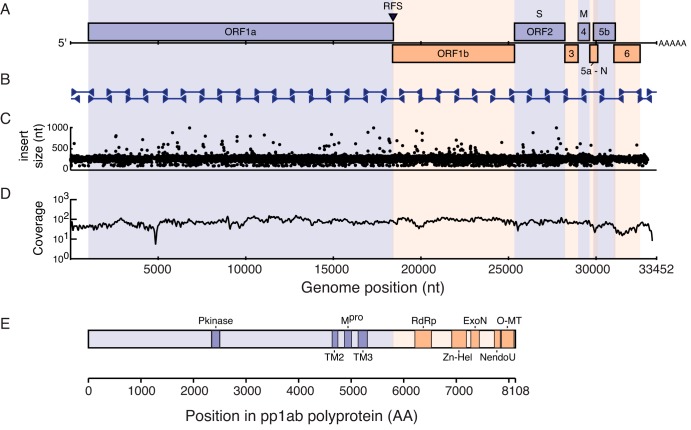
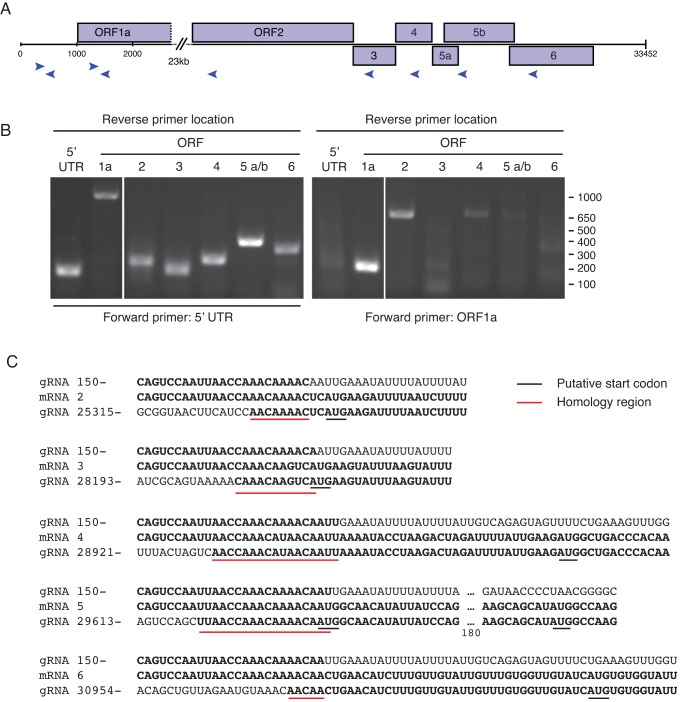
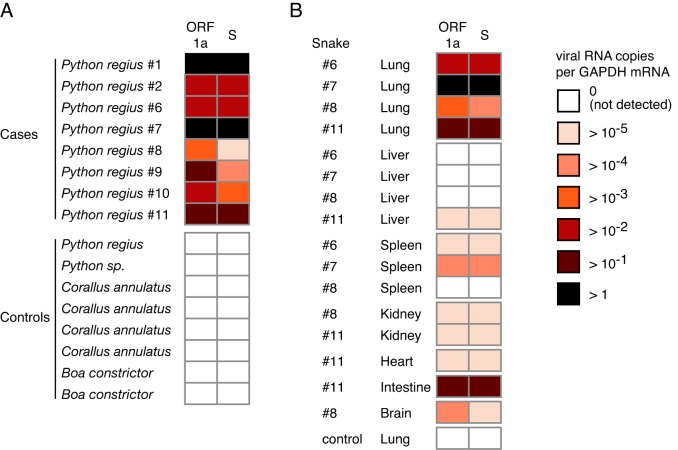
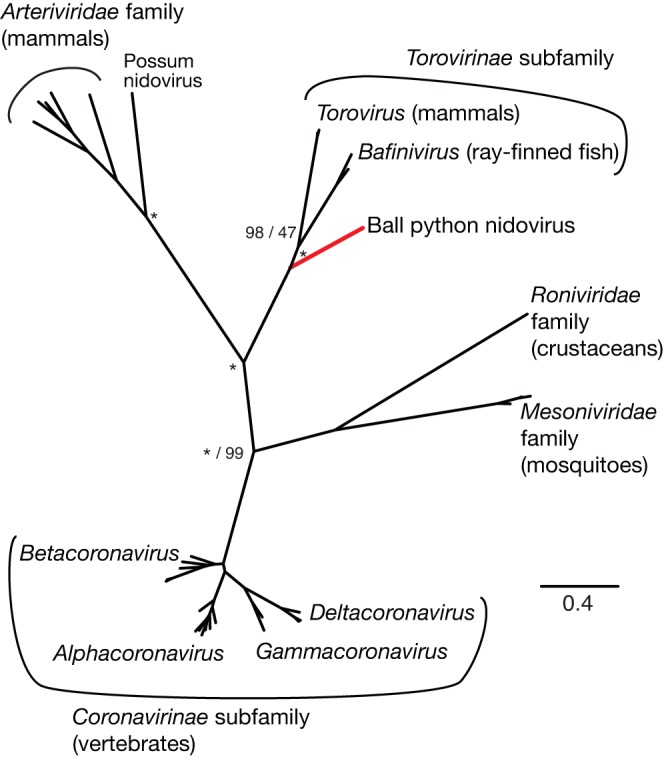
A severe, sometimes fatal respiratory disease has been observed in captive ball pythons (Python regius) since the late 1990s. In order to better understand this disease and its etiology, we collected case and control samples and performed pathological and diagnostic analyses. Electron micrographs revealed filamentous virus-like particles in lung epithelial cells of sick animals. Diagnostic testing for known pathogens did not identify an etiologic agent, so unbiased metagenomic sequencing was performed. Abundant nidovirus-like sequences were identified in cases and were used to assemble the genome of a previously unknown virus in the order Nidovirales. The nidoviruses, which were not previously known to infect nonavian reptiles, are a diverse order that includes important human and veterinary pathogens. The presence of the viral RNA was confirmed in all diseased animals (n = 8) but was not detected in healthy pythons or other snakes (n = 57). Viral RNA levels were generally highest in the lung and other respiratory tract tissues. The 33.5-kb viral genome is the largest RNA genome yet described and shares canonical characteristics with other nidovirus genomes, although several features distinguish this from related viruses. This virus, which we named ball python nidovirus (BPNV), will likely establish a new genus in Torovirinae subfamily. The identification of a novel nidovirus in reptiles contributes to our understanding of the biology and evolution of related viruses, and its association with lung disease in pythons is a promising step toward elucidating an etiology for this long-standing veterinary disease.
Importance: Ball pythons are popular pets because of their diverse coloration, generally nonaggressive behavior, and relatively small size. Since the 1990s, veterinarians have been aware of an infectious respiratory disease of unknown cause in ball pythons that can be fatal. We used unbiased shotgun sequencing to discover a novel virus in the order Nidovirales that was present in cases but not controls. While nidoviruses are known to infect a variety of animals, this is the first report of a nidovirus recovered from any reptile. This report will enable diagnostics that will assist in determining the role of this virus in the causation of disease, which would allow control of the disease in zoos and private collections. Given its evolutionary divergence from known nidoviruses and its unique host, the study of reptile nidoviruses may further our understanding of related diseases and the viruses that cause them in humans and other animals.
Copyright © 2014 Stenglein et al.
Figures


Similar articles
-
Investigations into the presence of nidoviruses in pythons.Virol J. 2020 Jan 17;17(1):6. doi: 10.1186/s12985-020-1279-5. Virol J. 2020. PMID: 31952524 Free PMC article.
-
Identification of a novel nidovirus in an outbreak of fatal respiratory disease in ball pythons (Python regius).Virol J. 2014 Aug 8;11:144. doi: 10.1186/1743-422X-11-144. Virol J. 2014. PMID: 25106433 Free PMC article.
-
Respiratory disease in ball pythons (Python regius) experimentally infected with ball python nidovirus.Virology. 2018 Apr;517:77-87. doi: 10.1016/j.virol.2017.12.008. Epub 2018 Jan 10. Virology. 2018. PMID: 29329683
-
Nidoviruses in Reptiles: A Review.Front Vet Sci. 2021 Sep 21;8:733404. doi: 10.3389/fvets.2021.733404. eCollection 2021. Front Vet Sci. 2021. PMID: 34621811 Free PMC article. Review.
-
Nidovirales: evolving the largest RNA virus genome.Virus Res. 2006 Apr;117(1):17-37. doi: 10.1016/j.virusres.2006.01.017. Epub 2006 Feb 28. Virus Res. 2006. PMID: 16503362 Free PMC article. Review.
Cited by
-
Characterization of Self-Processing Activities and Substrate Specificities of Porcine Torovirus 3C-Like Protease.J Virol. 2020 Sep 29;94(20):e01282-20. doi: 10.1128/JVI.01282-20. Print 2020 Sep 29. J Virol. 2020. PMID: 32727876 Free PMC article.
-
A fixed moderate-dose combination of tiletamine+zolazepam outperforms midazolam in induction of short-term immobilization of ball pythons (Python regius).PLoS One. 2018 Oct 19;13(10):e0199339. doi: 10.1371/journal.pone.0199339. eCollection 2018. PLoS One. 2018. PMID: 30339670 Free PMC article.
-
A case report of reptile-associated nidovirus (serpentovirus) in a ball python (Python regius) in Taiwan.J Vet Med Sci. 2020 Jun 24;82(6):788-792. doi: 10.1292/jvms.20-0166. Epub 2020 May 1. J Vet Med Sci. 2020. PMID: 32378646 Free PMC article.
-
Ebola virus, but not Marburg virus, replicates efficiently and without required adaptation in snake cells.Virus Evol. 2018 Nov 28;4(2):vey034. doi: 10.1093/ve/vey034. eCollection 2018 Jul. Virus Evol. 2018. PMID: 30524754 Free PMC article.
-
Identification and Characterization of Novel Serpentoviruses in Viperid and Elapid Snakes.Viruses. 2024 Sep 17;16(9):1477. doi: 10.3390/v16091477. Viruses. 2024. PMID: 39339954 Free PMC article.
References
-
- De Groot RJ, Cowley JA, Enjuanes L, Faaberg KS, Perlman S, Rottier PJM, Snijder EJ, Ziebuhr J, Gorbalenya AE. 2012. Nidovirales, p 785–795 In King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ. (ed), Virus taxonomy: classification and nomenclature of viruses: Ninth Report of the International Committee on Taxonomy of Viruses, 1st ed. Academic Press, Waltham, MA
-
- Lai MMC, Perlman S, Anderson LJ. 2007. Coronaviridae, p 1305–1335 In Knipe DM, Howley PM. (ed), Fields virology, 5th ed. Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Snijder EJ, Spaan WJM. 2007. Arteriviruses, p 1337–1355 In Knipe DM, Howley PM. (ed), Fields virology, 5th ed. Lippincott Williams & Wilkins, Philadelphia, PA.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
