

Reducing CD73 expression by IL1β-Programmed Th17 cells improves immunotherapeutic control of tumors
- PMID: 25205101
- PMCID: PMC4216762
- DOI: 10.1158/0008-5472.CAN-14-1450
Reducing CD73 expression by IL1β-Programmed Th17 cells improves immunotherapeutic control of tumors
Abstract
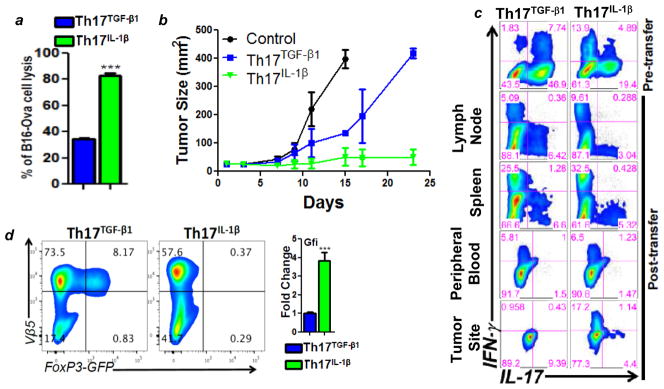
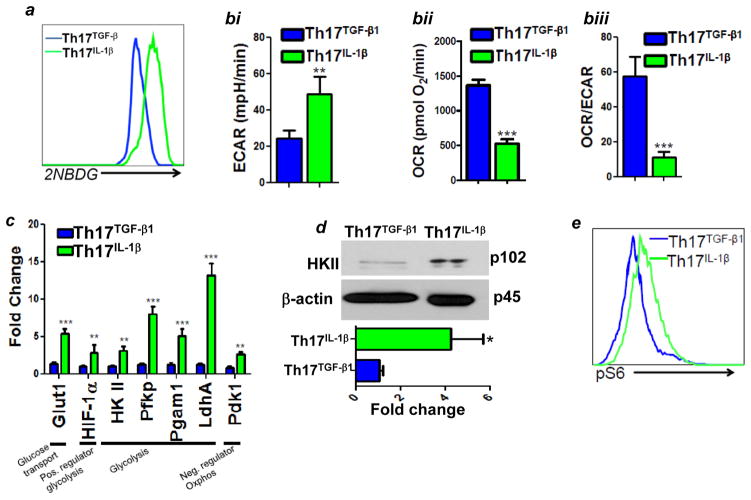
T cells of the T helper (Th)17 subset offer promise in adoptive T-cell therapy for cancer. However, current protocols for ex vivo programming of Th17 cells, which include TGFβ exposure, increase the expression of CD39 and CD73, two cell surface ATP ectonucleotidases that reduce T-cell effector functions and promote immunosuppression. Here, we report that ATP-mediated suppression of IFNγ production by Th17 cells can be overcome by genetic ablation of CD73 or by using IL1β instead of TGFβ to program Th17 cells ex vivo. Th17 cells cultured in IL1β were also highly polyfunctional, expressing high levels of effector molecules and exhibiting superior short-term control of melanoma in mice, despite reduced stem cell-like properties. TGFβ addition at low doses that did not upregulate CD73 expression but induced stemness properties drastically improved the antitumor effects of IL1β-cultured Th17 cells. Effector properties of IL1β-dependent Th17 cells were likely related to their high glycolytic capacity, since ex vivo programming in pyruvate impaired glycolysis and antitumor effects. Overall, we show that including TGFβ in ex vivo cultures used to program Th17 cells blunts their immunotherapeutic potential and demonstrate how this potential can be more fully realized for adoptive T-cell therapy.
©2014 American Association for Cancer Research.
Conflict of interest statement
Figures




References
-
- Ngiow SF, Teng MW, Smyth MJ. Prospects for TIM3-Targeted Antitumor Immunotherapy. Cancer Res. 2011;71(21):6567–71. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
