

MID1 catalyzes the ubiquitination of protein phosphatase 2A and mutations within its Bbox1 domain disrupt polyubiquitination of alpha4 but not of PP2Ac
- PMID: 25207814
- PMCID: PMC4160256
- DOI: 10.1371/journal.pone.0107428
MID1 catalyzes the ubiquitination of protein phosphatase 2A and mutations within its Bbox1 domain disrupt polyubiquitination of alpha4 but not of PP2Ac
Abstract
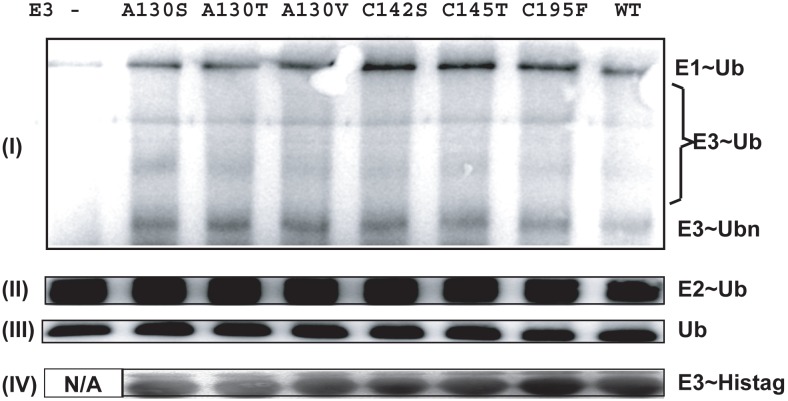
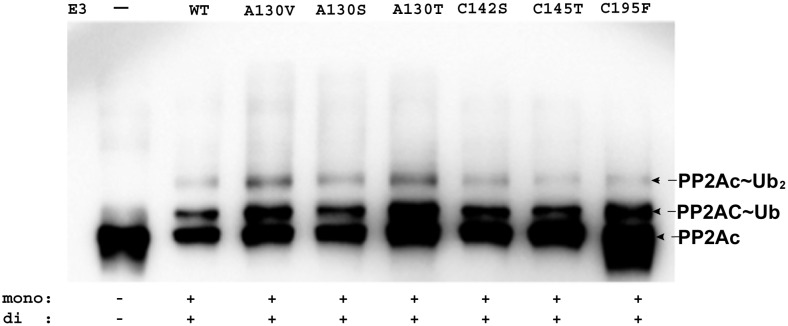
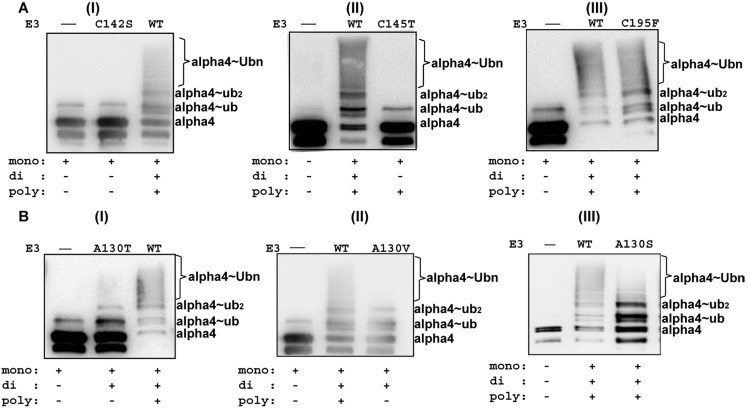
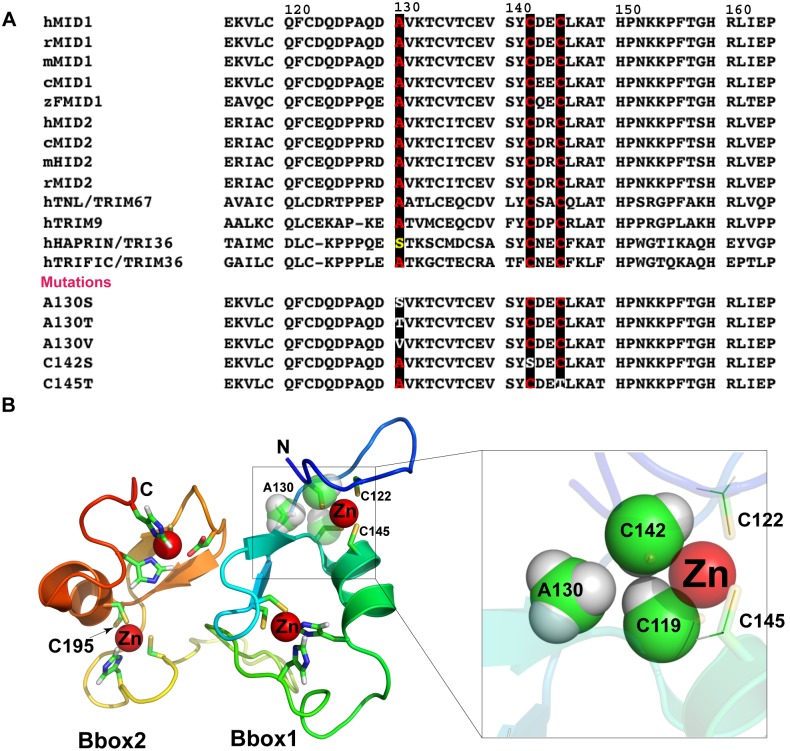
MID1 is a microtubule-associated protein that belongs to the TRIM family. MID1 functions as an ubiquitin E3 ligase, and recently was shown to catalyze the polyubiquitination of, alpha4, a protein regulator of protein phosphatase 2A (PP2A). It has been hypothesized that MID1 regulates PP2A, requiring the intermediary interaction with alpha4. Here we report that MID1 catalyzes the in vitro ubiquitination of the catalytic subunit of PP2A (PP2Ac) in the absence of alpha4. In the presence of alpha4, the level of PP2Ac ubiquitination is reduced. Using the MID1 RING-Bbox1-Bbox2 (RB1B2) construct containing the E3 ligase domains, we investigate the functional effects of mutations within the Bbox domains that are identified in patients with X-linked Opitz G syndrome (XLOS). The RB1B2 proteins harboring the C142S, C145T, A130V/T mutations within the Bbox1 domain and C195F mutation within the Bbox2 domain maintain auto-polyubiquitination activity. Qualitatively, the RB1B2 proteins containing these mutations are able to catalyze the ubiquitination of PP2Ac. In contrast, the RB1B2 proteins with mutations within the Bbox1 domain are unable to catalyze the polyubiquitination of alpha4. These results suggest that unregulated alpha4 may be the direct consequence of these natural mutations in the Bbox1 domain of MID1, and hence alpha4 could play a greater role to account for the increased amount of PP2A observed in XLOS-derived fibroblasts.
Conflict of interest statement
Figures


Similar articles
-
Structural and functional observations of the P151L MID1 mutation reveal alpha4 plays a significant role in X-linked Opitz Syndrome.FEBS J. 2017 Jul;284(14):2183-2193. doi: 10.1111/febs.14121. Epub 2017 Jun 14. FEBS J. 2017. PMID: 28548391
-
XLOS-observed mutations of MID1 Bbox1 domain cause domain unfolding.PLoS One. 2014 Sep 12;9(9):e107537. doi: 10.1371/journal.pone.0107537. eCollection 2014. PLoS One. 2014. PMID: 25216264 Free PMC article.
-
The E3 ubiquitin ligase MID1/TRIM18 promotes atypical ubiquitination of the BRCA2-associated factor 35, BRAF35.Biochim Biophys Acta Mol Cell Res. 2017 Oct;1864(10):1844-1854. doi: 10.1016/j.bbamcr.2017.07.014. Epub 2017 Jul 29. Biochim Biophys Acta Mol Cell Res. 2017. PMID: 28760657
-
The MID1 gene product in physiology and disease.Gene. 2020 Jul 15;747:144655. doi: 10.1016/j.gene.2020.144655. Epub 2020 Apr 10. Gene. 2020. PMID: 32283114 Free PMC article. Review.
-
X-linked Opitz syndrome: novel mutations in the MID1 gene and redefinition of the clinical spectrum.Am J Med Genet A. 2003 Jul 15;120A(2):222-8. doi: 10.1002/ajmg.a.10265. Am J Med Genet A. 2003. PMID: 12833403 Review.
Cited by
-
Molecular dynamics simulation reveals insights into the mechanism of unfolding by the A130T/V mutations within the MID1 zinc-binding Bbox1 domain.PLoS One. 2015 Apr 13;10(4):e0124377. doi: 10.1371/journal.pone.0124377. eCollection 2015. PLoS One. 2015. PMID: 25874572 Free PMC article.
-
Two Novel Pathogenic MID1 Variants and Genotype-Phenotype Correlation Reanalysis in X-Linked Opitz G/BBB Syndrome.Mol Syndromol. 2017 Dec;9(1):45-51. doi: 10.1159/000479177. Epub 2017 Aug 29. Mol Syndromol. 2017. PMID: 29456483 Free PMC article.
-
Mid1/Mid2 expression in craniofacial development and a literature review of X-linked opitz syndrome.Mol Genet Genomic Med. 2015 Dec 12;4(1):95-105. doi: 10.1002/mgg3.183. eCollection 2016 Jan. Mol Genet Genomic Med. 2015. PMID: 26788540 Free PMC article.
-
The ubiquitin E3 ligase NOSIP modulates protein phosphatase 2A activity in craniofacial development.PLoS One. 2014 Dec 29;9(12):e116150. doi: 10.1371/journal.pone.0116150. eCollection 2014. PLoS One. 2014. PMID: 25546391 Free PMC article.
-
TRIM Proteins and Antiviral Microtubule Reorganization: A Novel Component in Innate Immune Responses?Viruses. 2024 Aug 20;16(8):1328. doi: 10.3390/v16081328. Viruses. 2024. PMID: 39205302 Free PMC article. Review.
References
-
- Du H, Huang Y, Zaghlula M, Walters E, Cox TC, et al. (2013) The MID1 E3 Ligase Catalyzes the Polyubiquitination of Alpha4 (alpha4), a Regulatory Subunit of Protein Phosphatase 2A (PP2A): Novel Insights into MID1-mediated regulation of PP2A. The Journal of biological chemistry 288: 21341–21350. - PMC - PubMed
-
- Han X, Du H, Massiah MA (2011) Detection and Characterization of the In Vitro E3 Ligase Activity of the Human MID1 Protein. J Mol Biol 407: 505–520. - PubMed
-
- Kong M, Bui TV, Ditsworth D, Gruber JJ, Goncharov D, et al. (2007) The PP2A-associated protein alpha4 plays a critical role in the regulation of cell spreading and migration. The Journal of biological chemistry 282: 29712–29720. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
