

Time perception: the bad news and the good
- PMID: 25210578
- PMCID: PMC4142010
- DOI: 10.1002/wcs.1298
Time perception: the bad news and the good
Abstract
Time perception is fundamental and heavily researched, but the field faces a number of obstacles to theoretical progress. In this advanced review, we focus on three pieces of 'bad news' for time perception research: temporal perception is highly labile across changes in experimental context and task; there are pronounced individual differences not just in overall performance but in the use of different timing strategies and the effect of key variables; and laboratory studies typically bear little relation to timing in the 'real world'. We describe recent examples of these issues and in each case offer some 'good news' by showing how new research is addressing these challenges to provide rich insights into the neural and information-processing bases of timing and time perception. WIREs Cogn Sci 2014, 5:429-446. doi: 10.1002/wcs.1298 This article is categorized under: Psychology > Perception and Psychophysics Neuroscience > Cognition.
© 2014 The Authors. WIREs Cognitive Science published by John Wiley & Sons, Ltd.
Figures
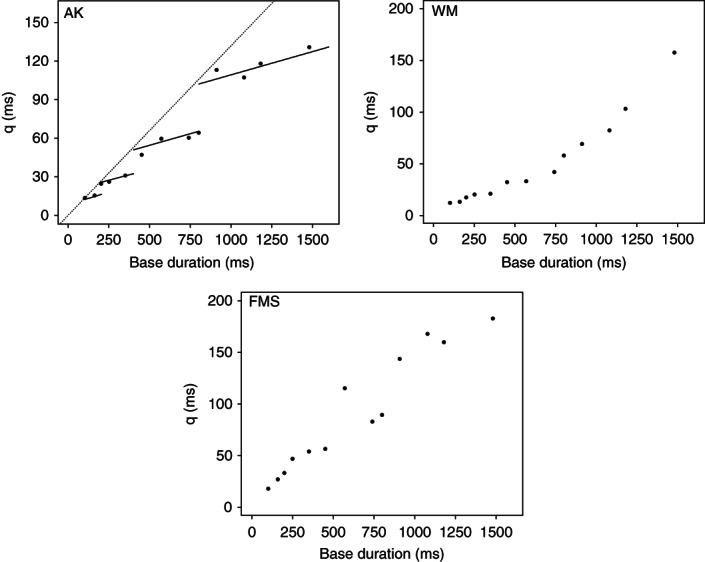
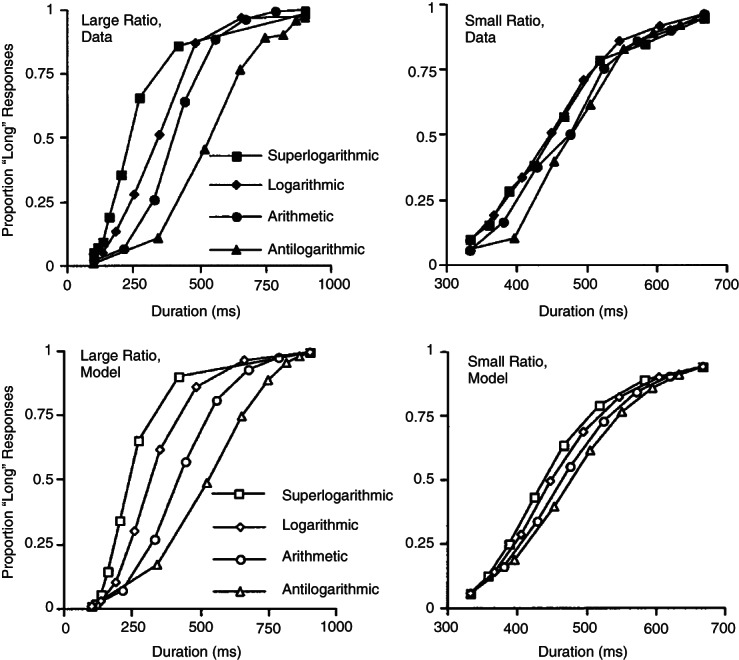
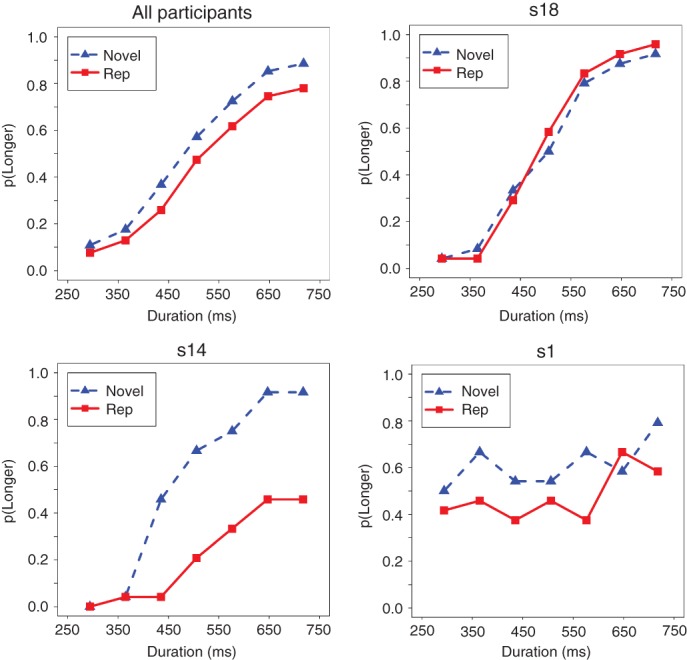
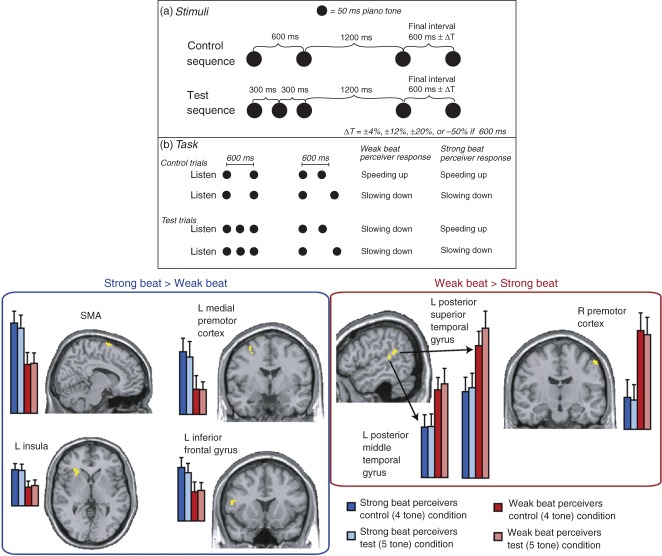
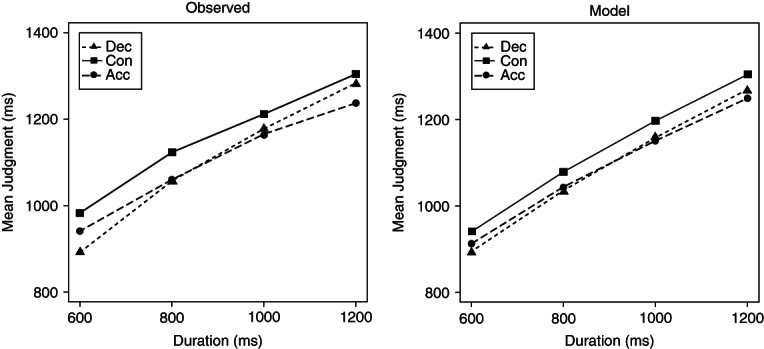
References
-
- Roeckelein JE. History of conceptions and accounts of time and early time perception research. In: Grondin S, editor. Psychology of Time. Bingley, UK: Emerald Press; 2008. pp. 1–50.
-
- James W. The Principles of Psychology. Vol. 1. New York: Holt; 1890. The perception of time.
-
- Wearden JH, Lejeune H. Scalar properties in human timing: conformity and violations. Q J Exp Psychol. 2008;61:569–587. - PubMed
-
- Lejeune H, Wearden JH. Scalar properties in animal timing: conformity and violations. Q J Exp Psychol. 2006;59:1875–1908. - PubMed
-
- Fechner GT. Elemente der psychophysik. Vol. 1. Leipzig: Breitkopf und Härtel; 1860.
FURTHER READING/RESOURCES
-
- Buhusi CV, Meck WH. What makes us tick? Functional and neural mechanisms of interval timing. Nat Rev Neurosci. 2005;6:755–765. - PubMed
-
- (These two p psychology and neurobiology of timing and time perception spanning a range from msec to 24 hrs in order to capture the interaction between circadian and interval timing mechanisms.)
-
- (Introduces mprising 58 research, opinion, and review articles covering multiple aspects of interval timing.)
RELATED ARTICLES
-
- Berniker M, Kording K. Bayesian approaches to sensory integration for motor control. WIREs Cognitive Sci. 2011;2:419–428. doi: 10.1002/wcs.125. - DOI - PubMed
-
- Braunlich K, Seger C. The basal ganglia. WIREs Cognitive Sci. 2013;4:135–148. doi: 10.1002/wcs.1217. - DOI - PubMed
-
- Goel V. Neural basis of thinking: laboratory problems versus real-world problems. WIREs Cognitive Sci. 2010;1:613–621. doi: 10.1002/wcs.71. - DOI - PubMed
-
- Sasaki Y, Náñez JE, Watanabe T. Recent progress in perceptual learning research. WIREs Cognitive Sci. 2012;3:293–299. doi: 10.1002/wcs.1175. - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
