

Strong epistatic selection on the RNA secondary structure of HIV
- PMID: 25210786
- PMCID: PMC4161434
- DOI: 10.1371/journal.ppat.1004363
Strong epistatic selection on the RNA secondary structure of HIV
Abstract
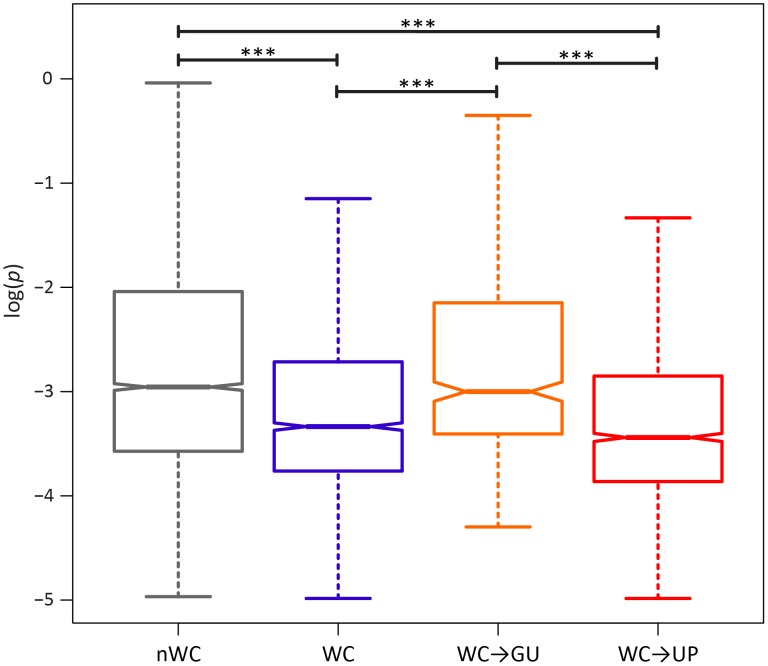
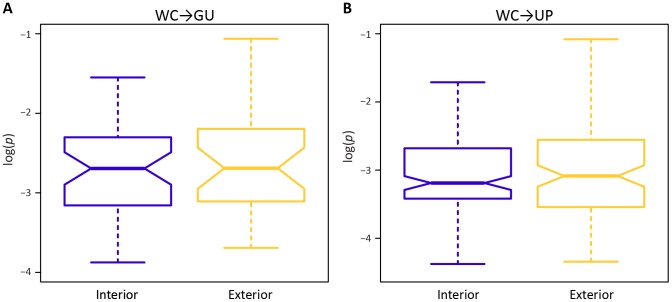
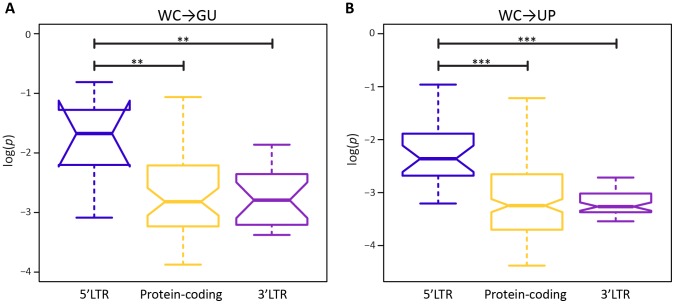
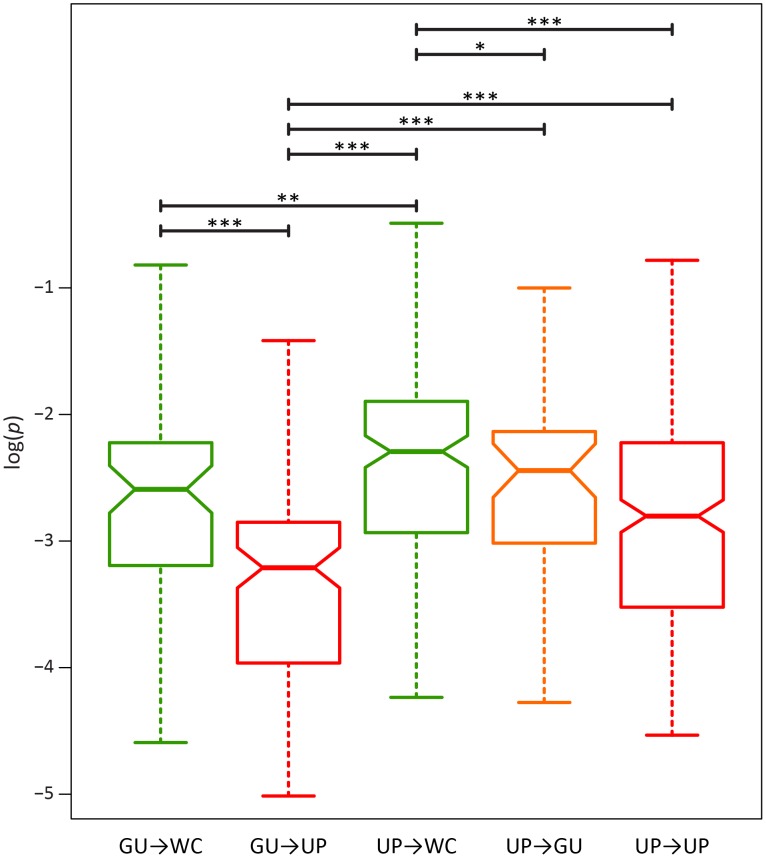
A key question in evolutionary genomics is how populations navigate the adaptive landscape in the presence of epistasis, or interactions among loci. This problem can be directly addressed by studying the evolution of RNA secondary structures, for which there is constraint to maintain pairing between Watson-Crick (WC) sites. Replacement of a nucleotide at one site of a WC pair reduces fitness by disrupting binding, which can be restored via a compensatory replacement at the interacting site. Here, I present the first genome-scale analysis of epistasis on the RNA secondary structure of human immunodeficiency virus type 1 (HIV-1). Comparison of polymorphism frequencies at ancestrally conserved sites reveals that selection against replacements is ∼ 2.7 times stronger at WC than at non-WC sites, such that nearly 50% of constraint can be attributed to epistasis. However, almost all epistatic constraint is due to selection against conversions of WC pairs to unpaired (UP) nucleotides, whereas conversions to GU wobbles are only slightly deleterious. This disparity is also evident in pairs with second-site compensatory replacements; conversions from UP nucleotides to WC pairs increase median fitness by ∼ 4.2%, whereas conversions from GU wobbles to WC pairs only increase median fitness by ∼ 0.3%. Moreover, second-site replacements that convert UP nucleotides to GU wobbles also increase median fitness by ∼ 4%, indicating that such replacements are nearly as compensatory as those that restore WC pairing. Thus, WC peaks of the HIV-1 epistatic adaptive landscape are connected by high GU ridges, enabling the viral population to rapidly explore distant peaks without traversing deep UP valleys.
Conflict of interest statement
The author has declared that no competing interests exist.
Figures
References
-
- Kimura M (1985) The role of compensatory neutral mutations in molecular evolution. J Genet 64: 7–19.
-
- Huynen MA, Hogeweg P (1994) Pattern generation in molecular evolution: exploitation of the variation in RNA landscapes. J Mol Evol 39: 71–79. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
