

Translational regulation of the DOUBLETIME/CKIδ/ε kinase by LARK contributes to circadian period modulation
- PMID: 25211129
- PMCID: PMC4161311
- DOI: 10.1371/journal.pgen.1004536
Translational regulation of the DOUBLETIME/CKIδ/ε kinase by LARK contributes to circadian period modulation
Abstract
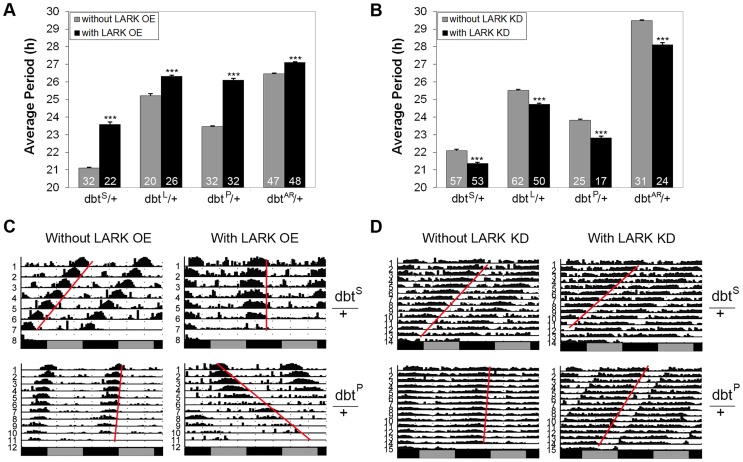
The Drosophila homolog of Casein Kinase I δ/ε, DOUBLETIME (DBT), is required for Wnt, Hedgehog, Fat and Hippo signaling as well as circadian clock function. Extensive studies have established a critical role of DBT in circadian period determination. However, how DBT expression is regulated remains largely unexplored. In this study, we show that translation of dbt transcripts are directly regulated by a rhythmic RNA-binding protein (RBP) called LARK (known as RBM4 in mammals). LARK promotes translation of specific alternative dbt transcripts in clock cells, in particular the dbt-RC transcript. Translation of dbt-RC exhibits circadian changes under free-running conditions, indicative of clock regulation. Translation of a newly identified transcript, dbt-RE, is induced by light in a LARK-dependent manner and oscillates under light/dark conditions. Altered LARK abundance affects circadian period length, and this phenotype can be modified by different dbt alleles. Increased LARK delays nuclear degradation of the PERIOD (PER) clock protein at the beginning of subjective day, consistent with the known role of DBT in PER dynamics. Taken together, these data support the idea that LARK influences circadian period and perhaps responses of the clock to light via the regulated translation of DBT. Our study is the first to investigate translational control of the DBT kinase, revealing its regulation by LARK and a novel role of this RBP in Drosophila circadian period modulation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







Comment in
-
Translational regulation of the Drosophila post-translational circadian mechanism.PLoS Genet. 2014 Sep 11;10(9):e1004628. doi: 10.1371/journal.pgen.1004628. eCollection 2014 Sep. PLoS Genet. 2014. PMID: 25210865 Free PMC article. No abstract available.
References
-
- Kloss B, Price JL, Saez L, Blau J, Rothenfluh A, et al. (1998) The Drosophila clock gene double-time encodes a protein closely related to human casein kinase Iepsilon. Cell 94: 97–107. - PubMed
-
- Price JL, Blau J, Rothenfluh A, Abodeely M, Kloss B, et al. (1998) double-time is a novel Drosophila clock gene that regulates PERIOD protein accumulation. Cell 94: 83–95. - PubMed
-
- Zilian O, Frei E, Burke R, Brentrup D, Gutjahr T, et al. (1999) double-time is identical to discs overgrown, which is required for cell survival, proliferation and growth arrest in Drosophila imaginal discs. Development 126: 5409–5420. - PubMed
-
- Klein TJ, Jenny A, Djiane A, Mlodzik M (2006) CKIepsilon/discs overgrown promotes both Wnt-Fz/beta-catenin and Fz/PCP signaling in Drosophila. Curr Biol 16: 1337–1343. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
