

Bacillithiol is a major buffer of the labile zinc pool in Bacillus subtilis
- PMID: 25213752
- PMCID: PMC4227968
- DOI: 10.1111/mmi.12794
Bacillithiol is a major buffer of the labile zinc pool in Bacillus subtilis
Abstract
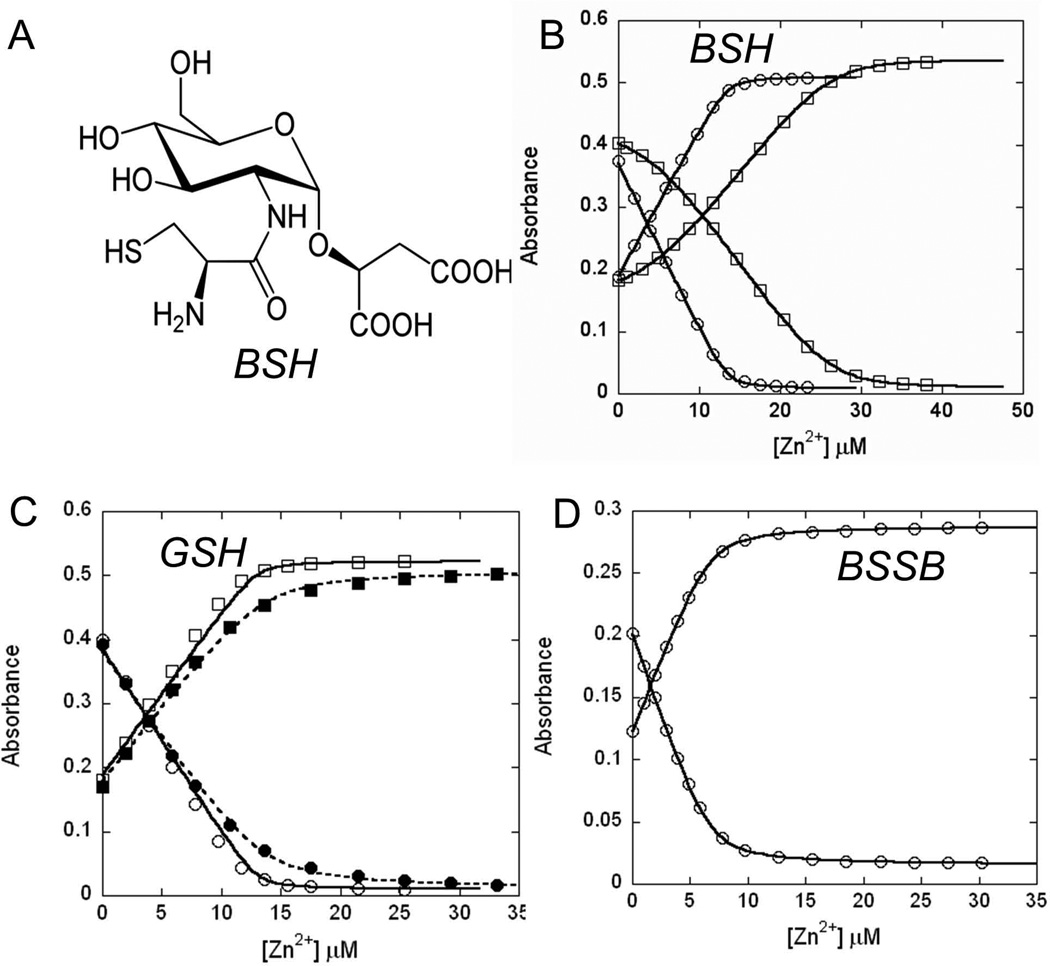
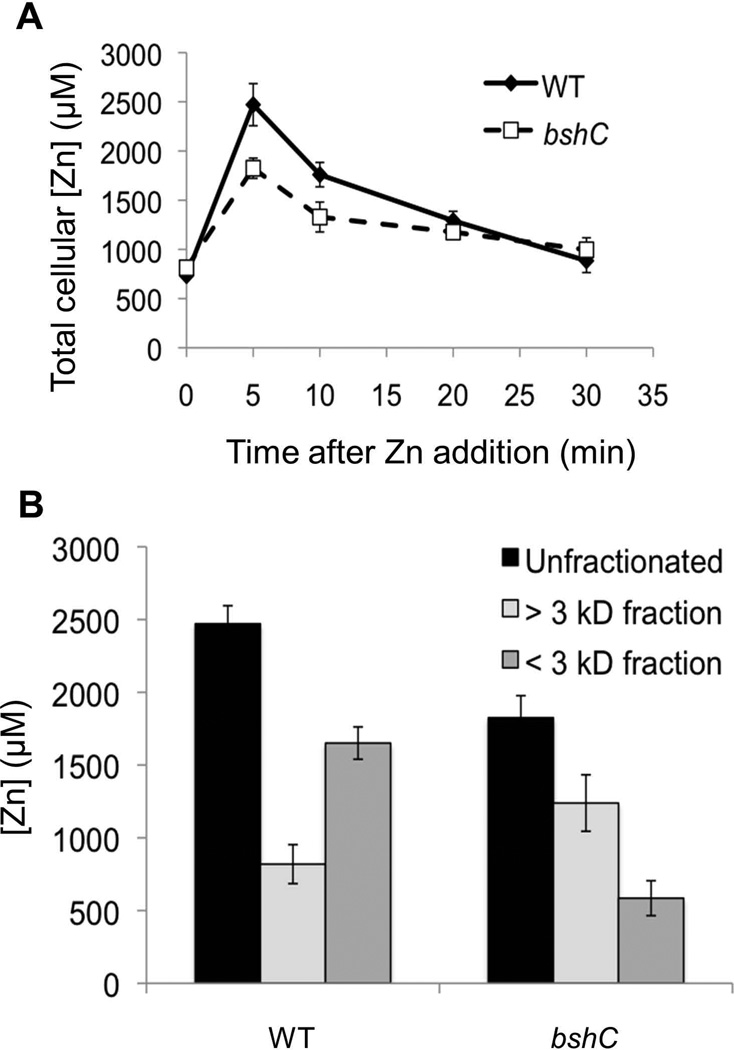
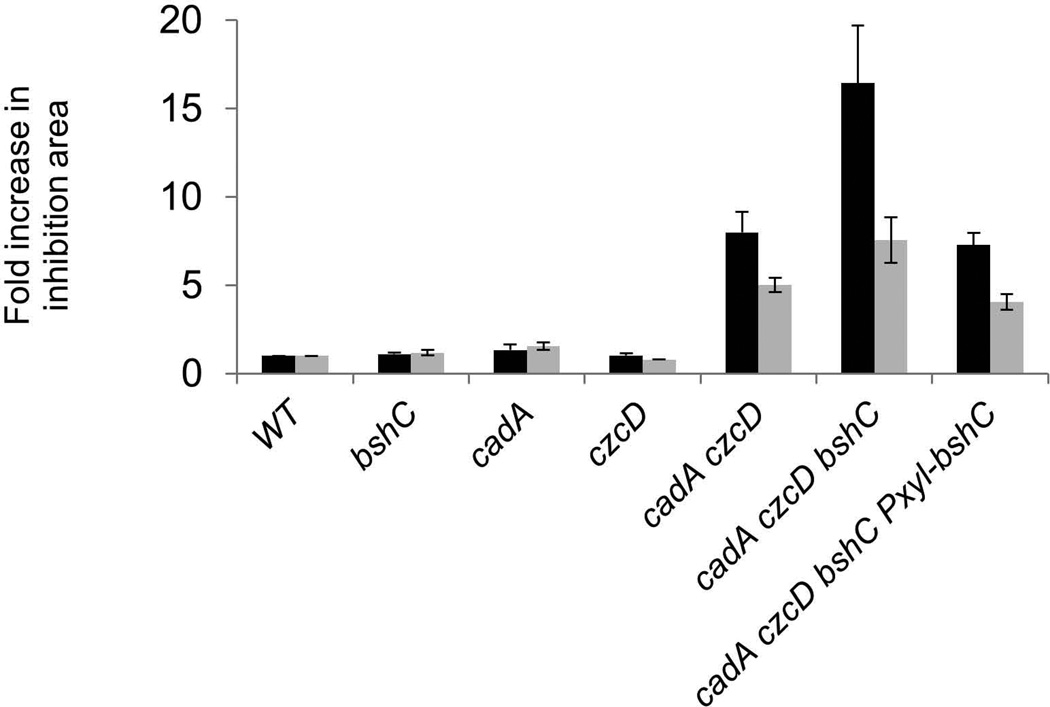
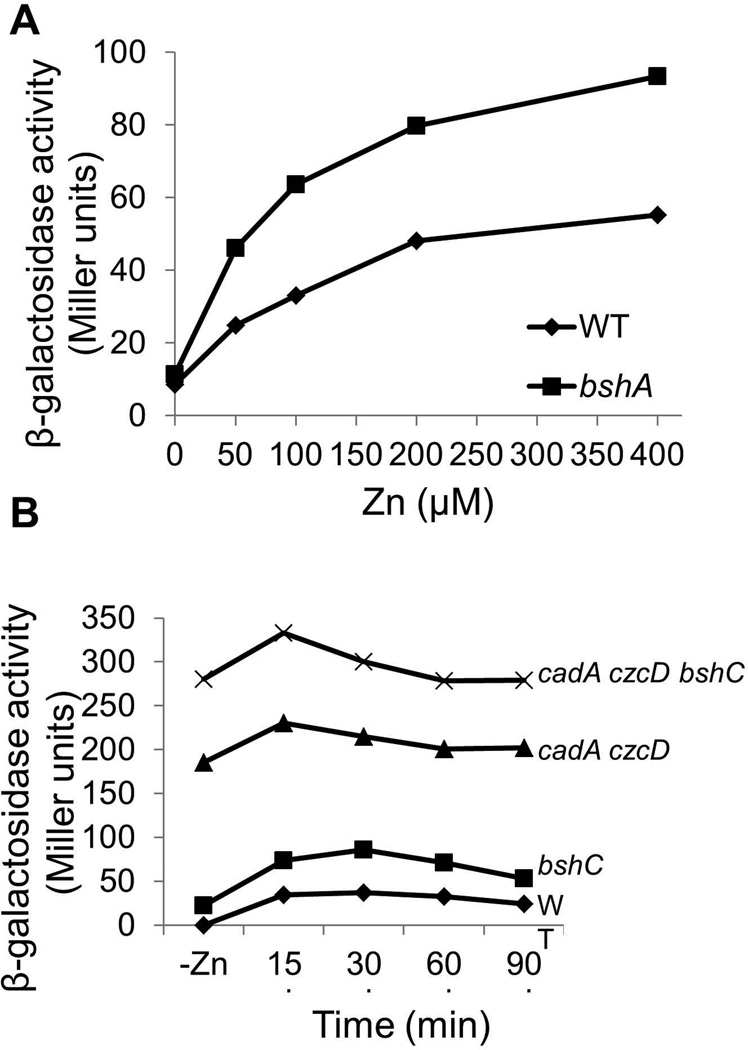
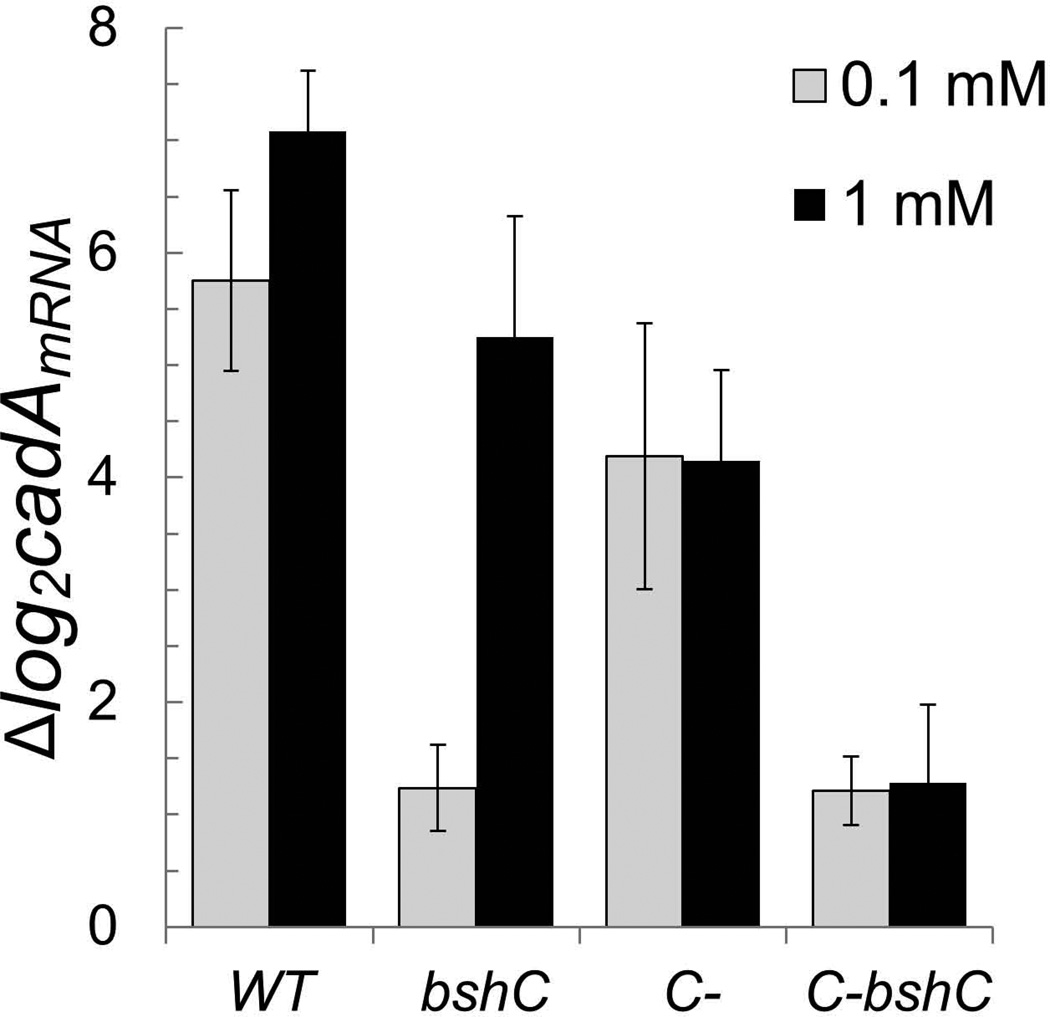
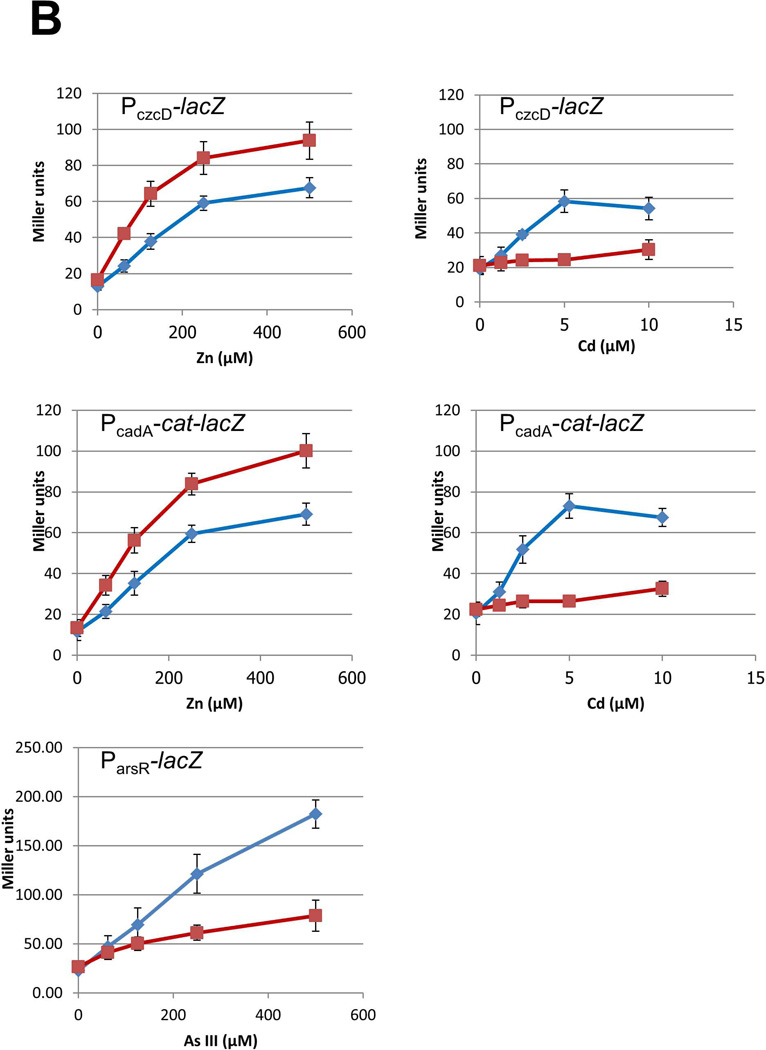
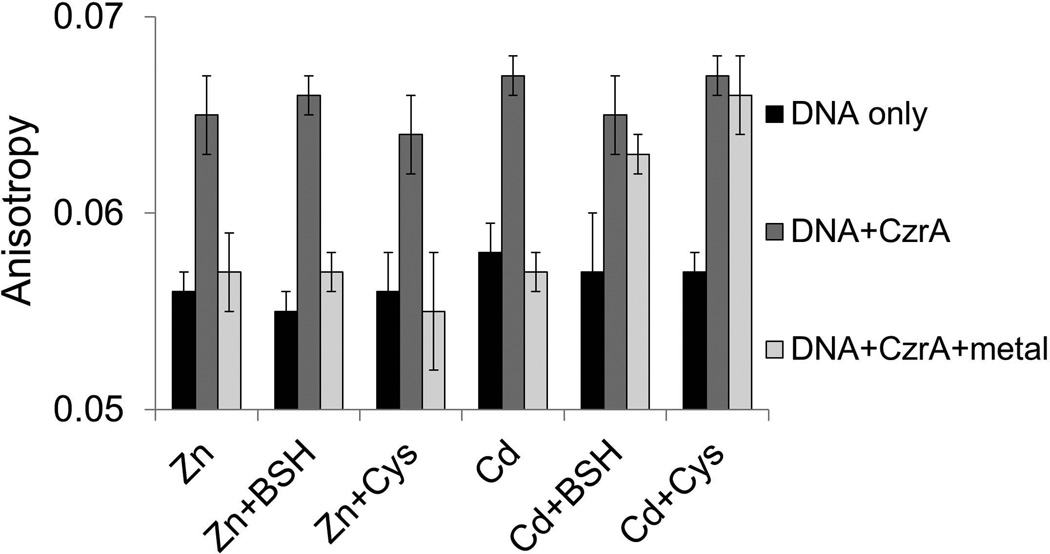
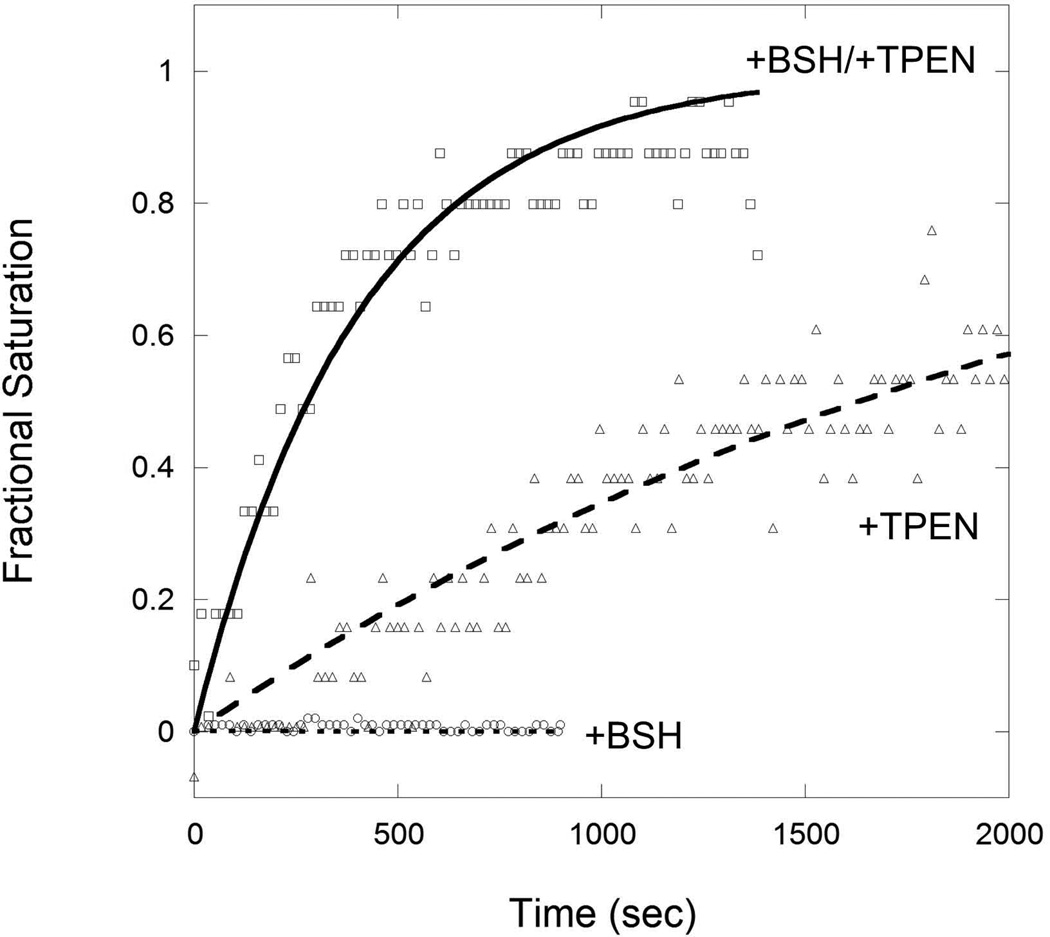
Intracellular zinc levels are tightly regulated since zinc is an essential cofactor for numerous enzymes, yet can be toxic when present in excess. The majority of intracellular zinc is tightly associated with proteins and is incorporated during synthesis from a poorly defined pool of kinetically labile zinc. In Bacillus subtilis, this labile pool is sensed by equilibration with the metalloregulator Zur, as an indication of zinc sufficiency, and by CzrA, as an indication of zinc excess. Here, we demonstrate that the low-molecular-weight thiol bacillithiol (BSH) serves as a major buffer of the labile zinc pool. Upon shift to conditions of zinc excess, cells transiently accumulate zinc in a low-molecular-weight pool, and this accumulation is largely dependent on BSH. Cells lacking BSH are more sensitive to zinc stress, and they induce zinc efflux at lower external zinc concentrations. Thiol reactive agents such as diamide and cadmium induce zinc efflux by interfering with the Zn-buffering function of BSH. Our data provide new insights into intracellular zinc buffering and may have broad relevance given the presence of BSH in pathogens and the proposed role of zinc sequestration in innate immunity.
© 2014 John Wiley & Sons Ltd.
Figures
Comment in
-
Bacillithiol, a new role in buffering intracellular zinc.Mol Microbiol. 2014 Nov;94(4):743-6. doi: 10.1111/mmi.12793. Epub 2014 Sep 30. Mol Microbiol. 2014. PMID: 25213645 Free PMC article.
References
-
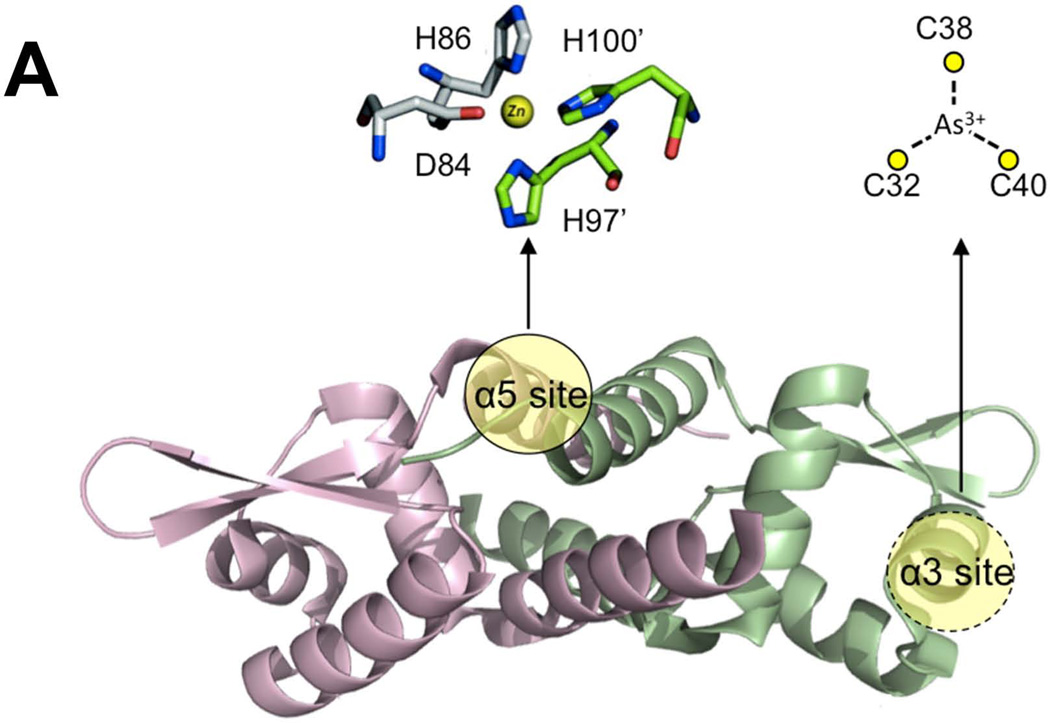
- Busenlehner LS, Pennella MA, Giedroc DP. The SmtB/ArsR family of metalloregulatory transcriptional repressors: Structural insights into prokaryotic metal resistance. FEMS microbiology reviews. 2003;27:131–143. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
