

Resetting transcription factor control circuitry toward ground-state pluripotency in human
- PMID: 25215486
- PMCID: PMC4162745
- DOI: 10.1016/j.cell.2014.08.029
Resetting transcription factor control circuitry toward ground-state pluripotency in human
Erratum in
- Cell. 2015 Jul 16;162(2):152-3
-
Resetting Transcription Factor Control Circuitry toward Ground-State Pluripotency in Human.Cell. 2015 Jul 16;162(2):452-453. doi: 10.1016/j.cell.2015.06.052. Epub 2015 Jul 16. Cell. 2015. PMID: 28843285 Free PMC article. No abstract available.
Abstract
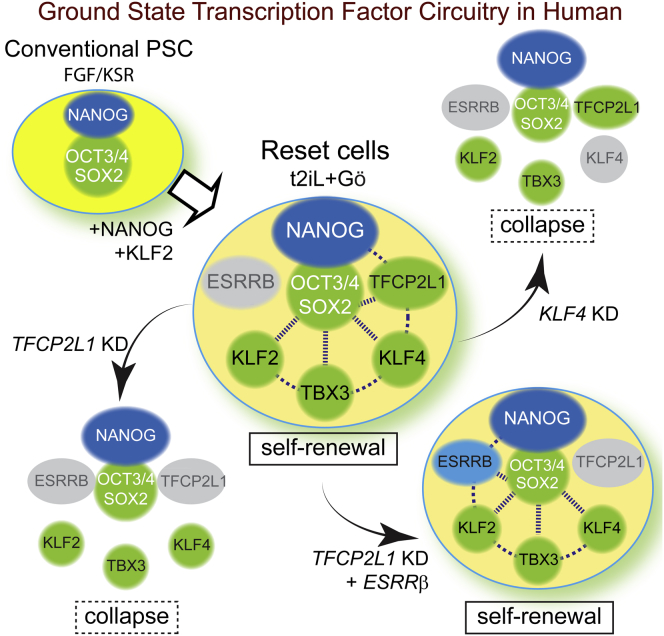
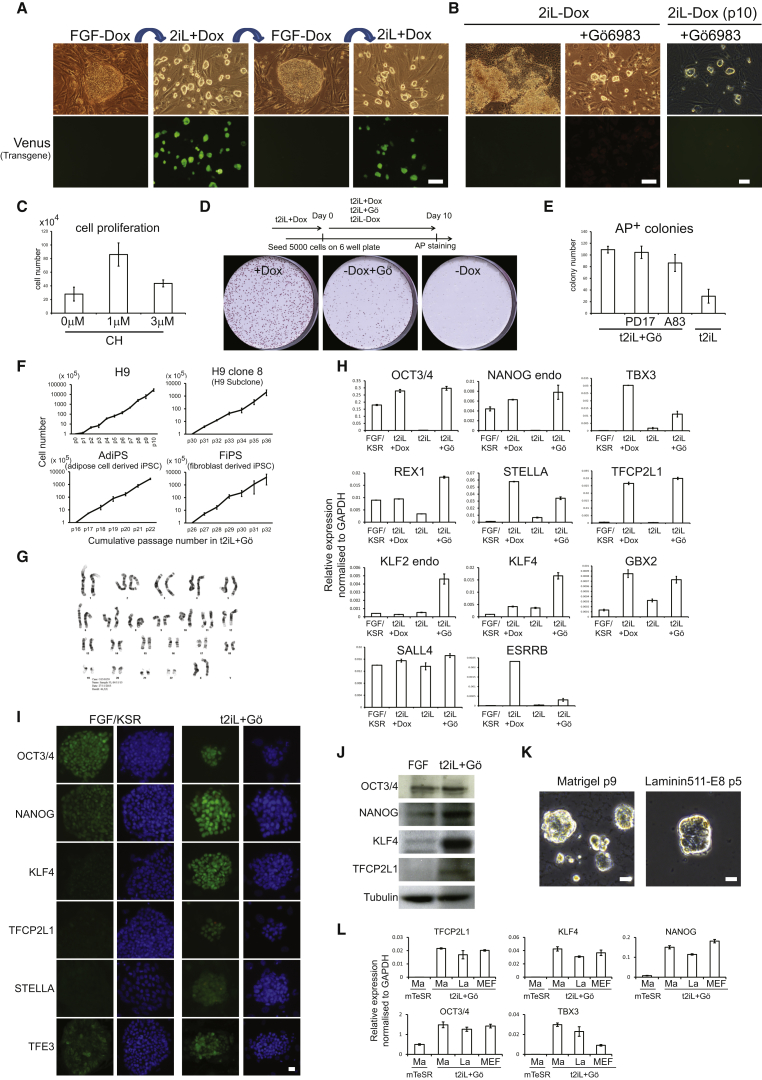
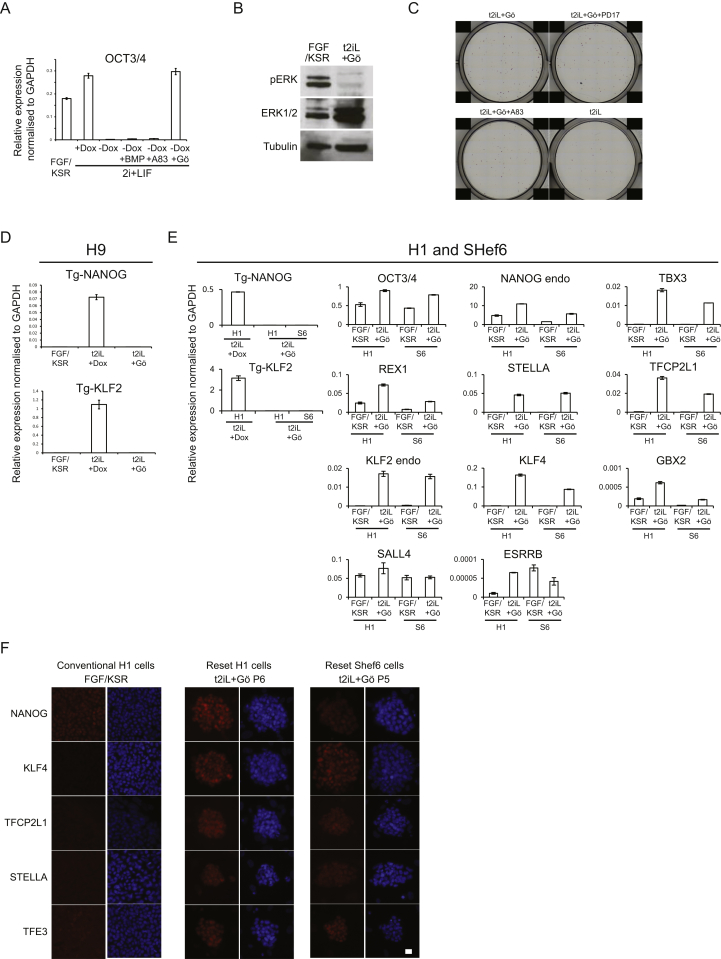
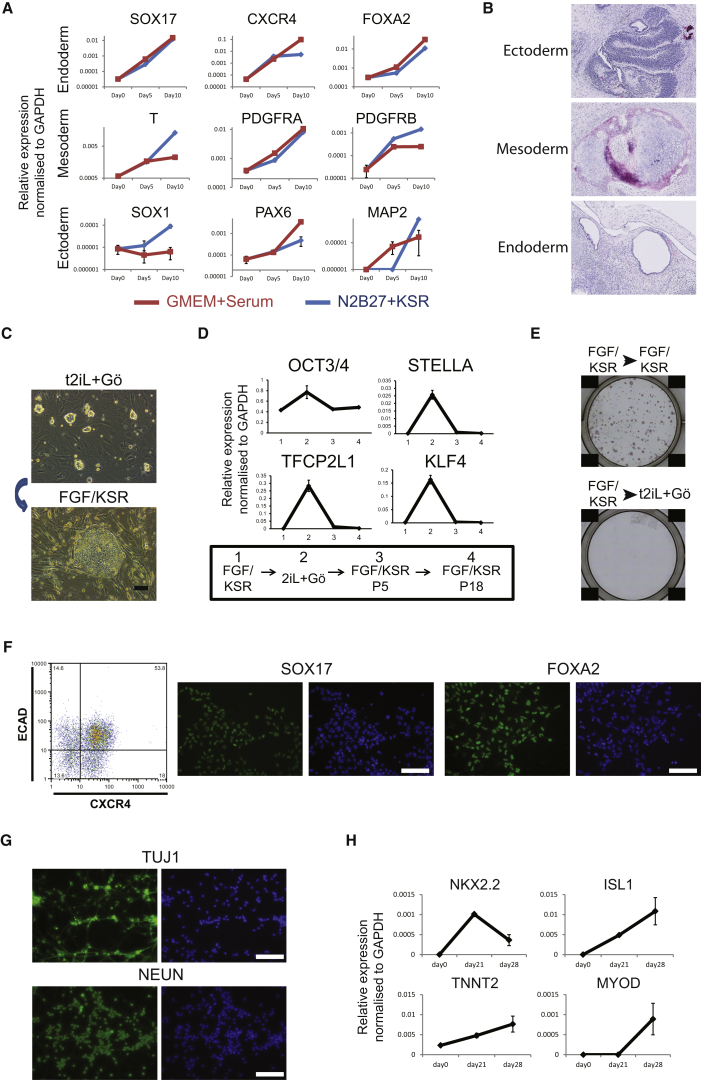
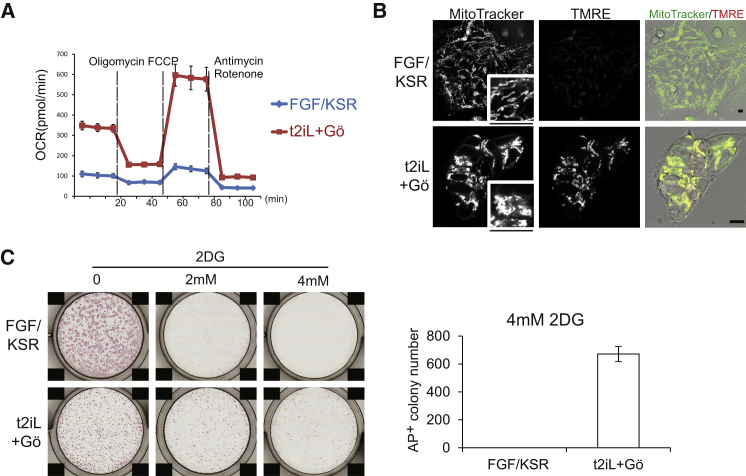
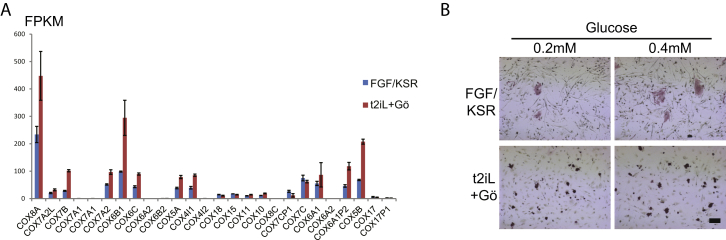
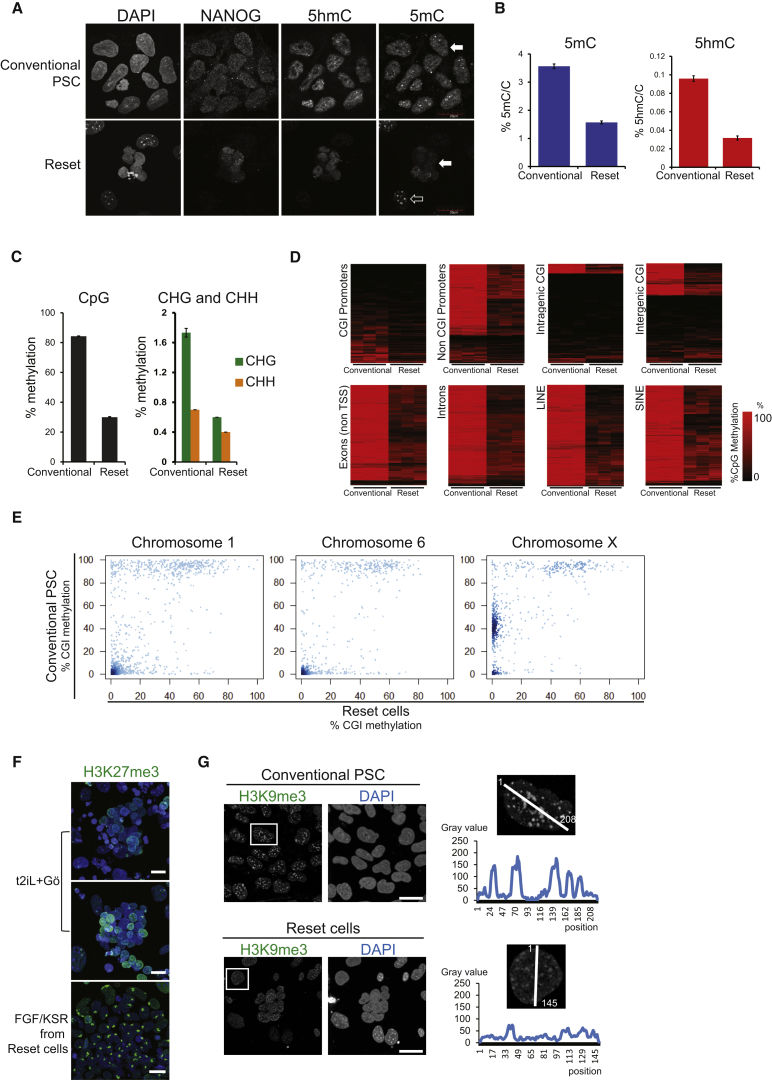
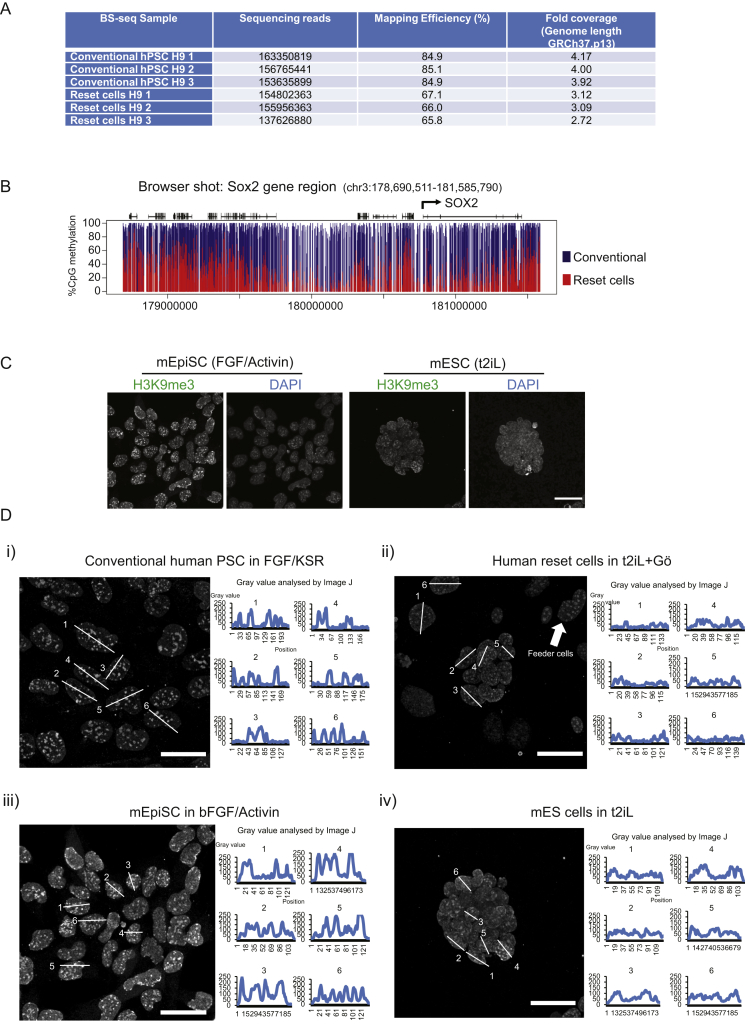
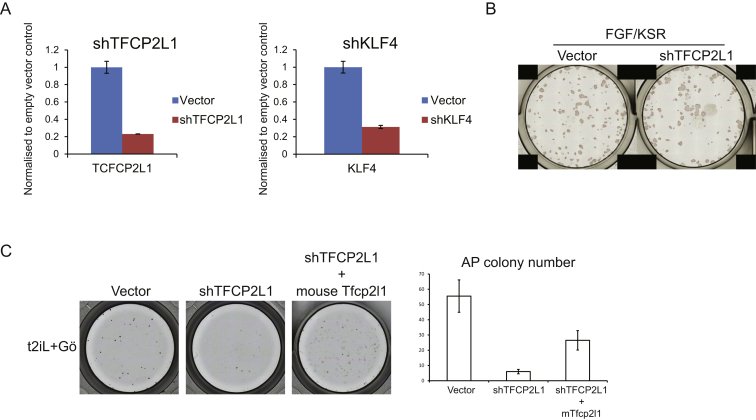
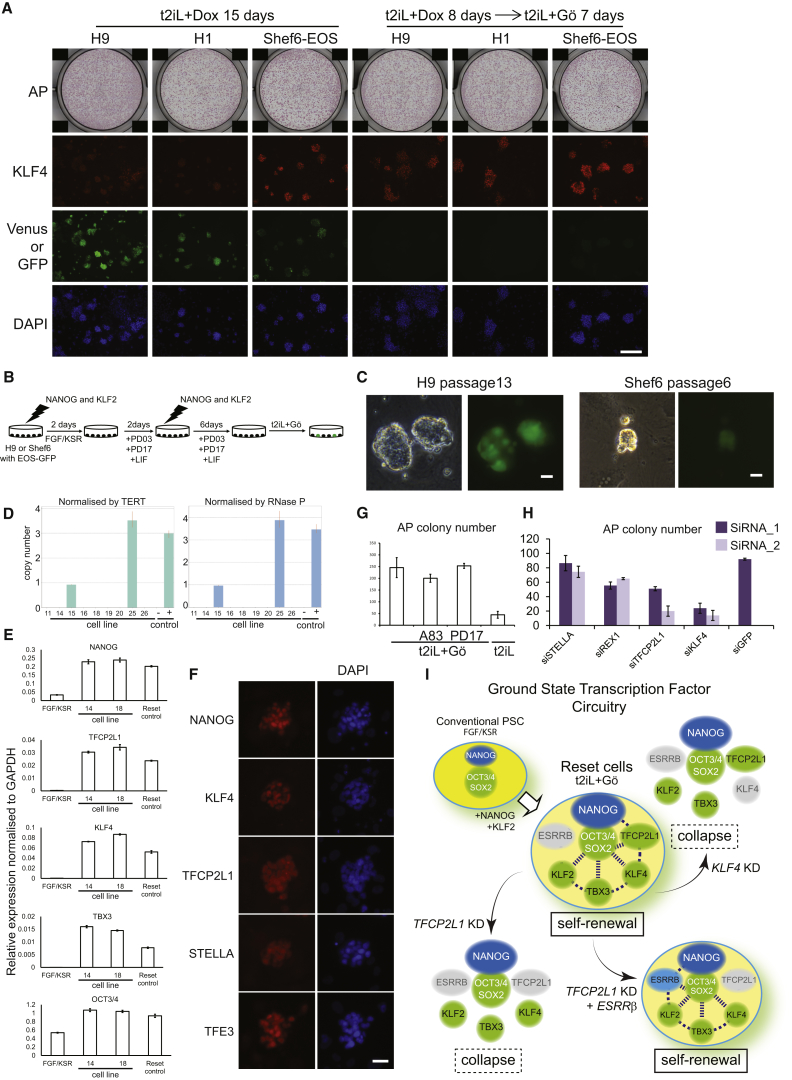
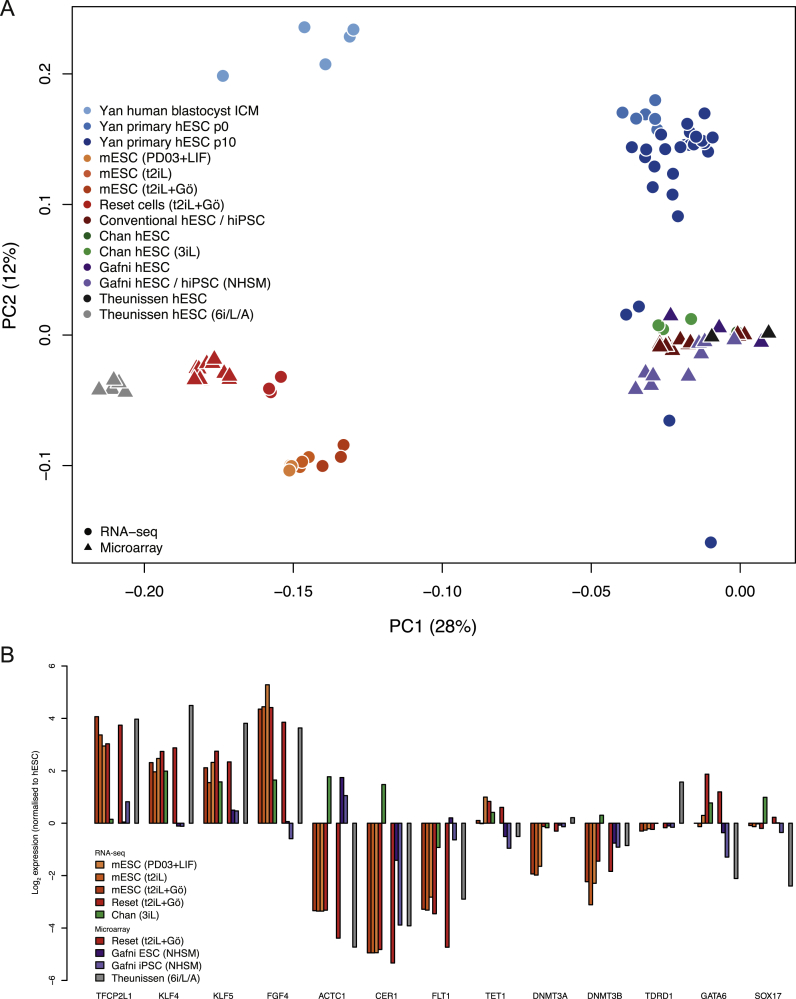
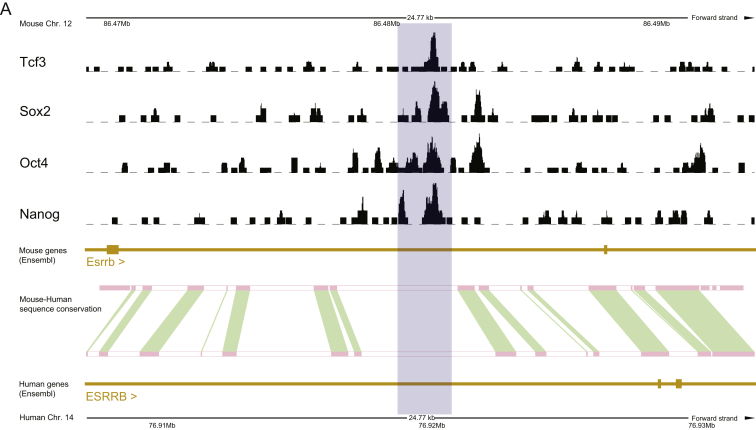
Current human pluripotent stem cells lack the transcription factor circuitry that governs the ground state of mouse embryonic stem cells (ESC). Here, we report that short-term expression of two components, NANOG and KLF2, is sufficient to ignite other elements of the network and reset the human pluripotent state. Inhibition of ERK and protein kinase C sustains a transgene-independent rewired state. Reset cells self-renew continuously without ERK signaling, are phenotypically stable, and are karyotypically intact. They differentiate in vitro and form teratomas in vivo. Metabolism is reprogrammed with activation of mitochondrial respiration as in ESC. DNA methylation is dramatically reduced and transcriptome state is globally realigned across multiple cell lines. Depletion of ground-state transcription factors, TFCP2L1 or KLF4, has marginal impact on conventional human pluripotent stem cells but collapses the reset state. These findings demonstrate feasibility of installing and propagating functional control circuitry for ground-state pluripotency in human cells.
Copyright © 2014 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
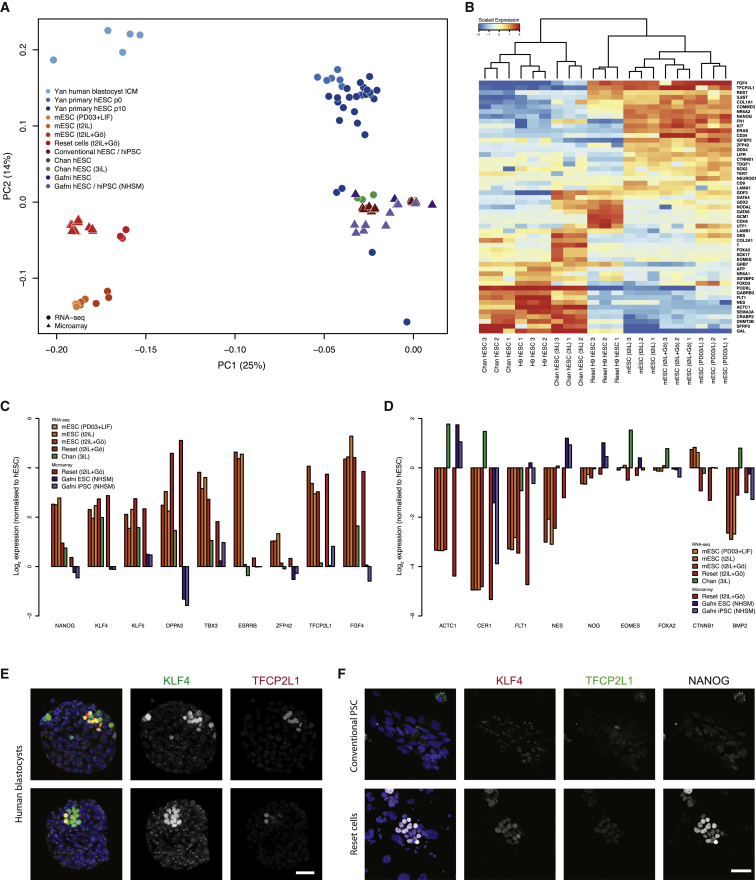
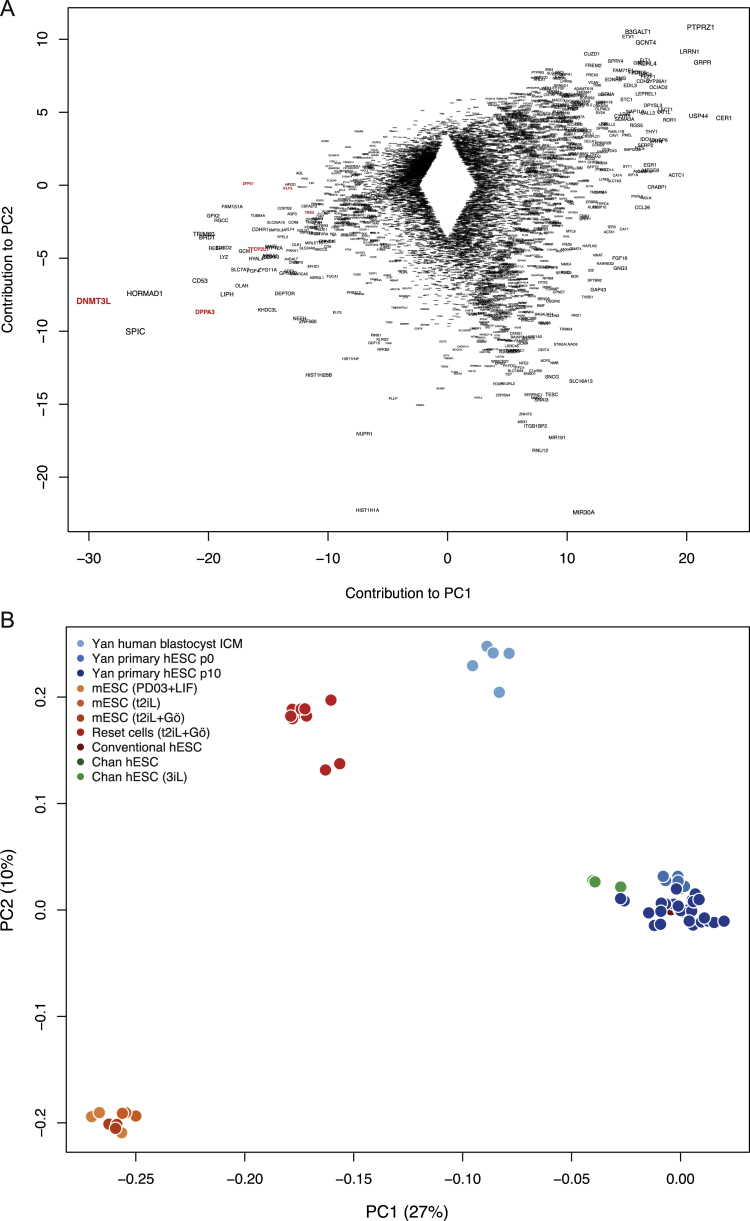
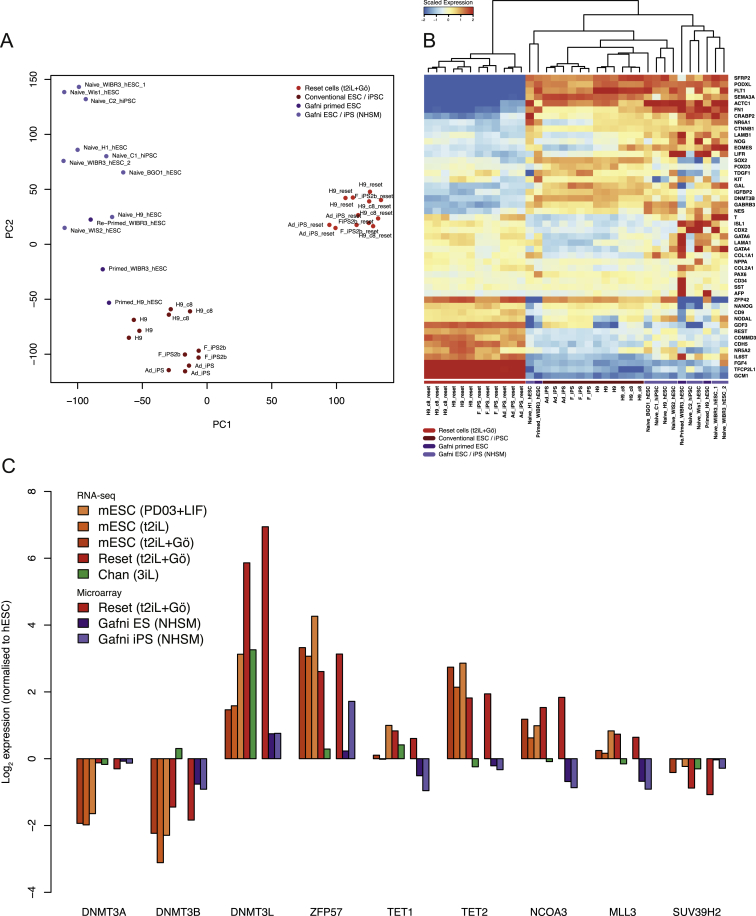
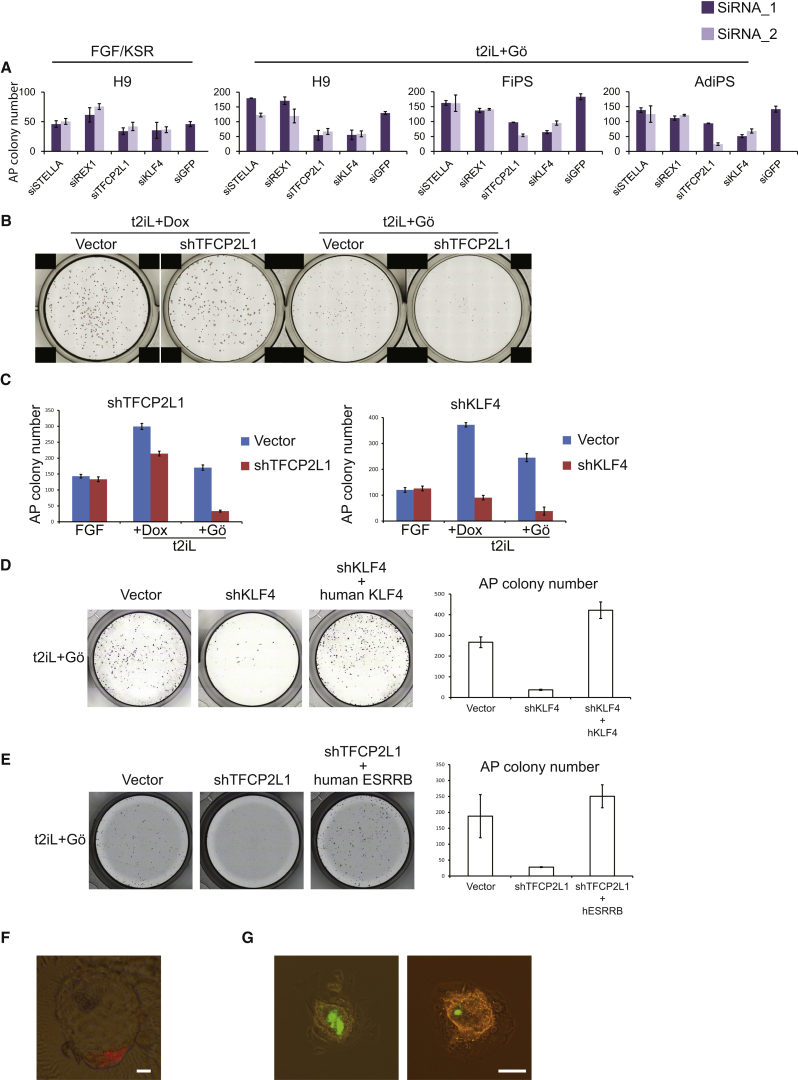
References
-
- Adewumi O., Aflatoonian B., Ahrlund-Richter L., Amit M., Andrews P.W., Beighton G., Bello P.A., Benvenisty N., Berry L.S., Bevan S., International Stem Cell Initiative Characterization of human embryonic stem cell lines by the International Stem Cell Initiative. Nat. Biotechnol. 2007;25:803–816. - PubMed
-
- Amit M., Carpenter M.K., Inokuma M.S., Chiu C.P., Harris C.P., Waknitz M.A., Itskovitz-Eldor J., Thomson J.A. Clonally derived human embryonic stem cell lines maintain pluripotency and proliferative potential for prolonged periods of culture. Dev. Biol. 2000;227:271–278. - PubMed
Supplemental References
-
- Aflatoonian B., Ruban L., Shamsuddin S., Baker D., Andrews P., Moore H. Generation of Sheffield (Shef) human embryonic stem cell lines using a microdrop culture system. In Vitro Cell. Dev. Biol. Anim. 2010;46:236–241. - PubMed
-
- Moretti A., Bellin M., Jung C.B., Thies T.M., Takashima Y., Bernshausen A., Schiemann M., Fischer S., Moosmang S., Smith A.G. Mouse and human induced pluripotent stem cells as a source for multipotent Isl1+ cardiovascular progenitors. FASEB J. 2010;24:700–711. - PubMed
-
- Thomson J.A., Itskovitz-Eldor J., Shapiro S.S., Waknitz M.A., Swiergiel J.J., Marshall V.S., Jones J.M. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–1147. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
