

Forced IDO1 expression in dendritic cells restores immunoregulatory signalling in autoimmune diabetes
- PMID: 25215657
- PMCID: PMC4193887
- DOI: 10.1111/jcmm.12360
Forced IDO1 expression in dendritic cells restores immunoregulatory signalling in autoimmune diabetes
Abstract
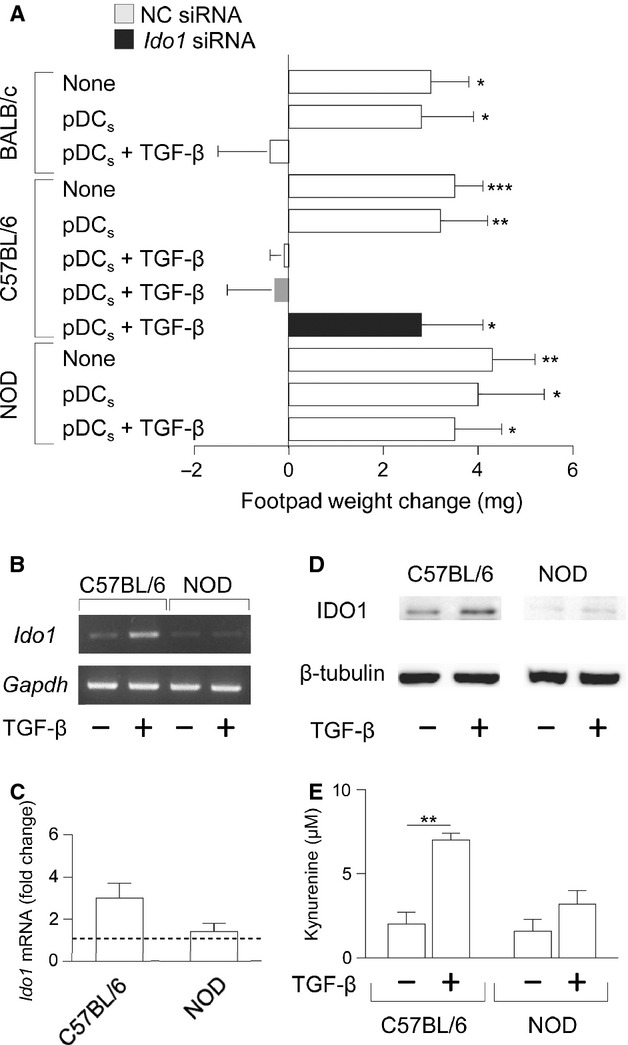
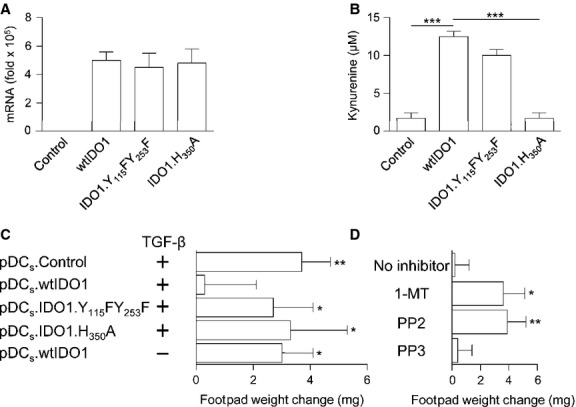
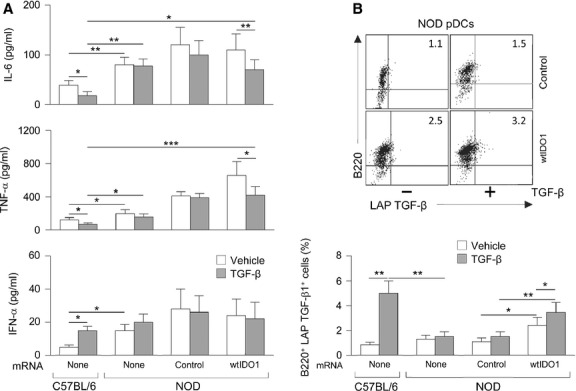
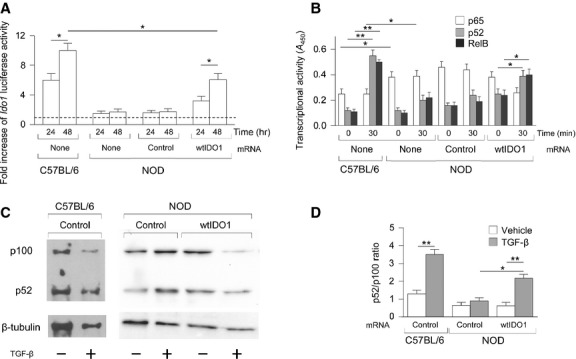
Indoleamine 2,3-dioxygenase (IDO1), a tryptophan catabolizing enzyme, is recognized as an authentic regulator of immunity in several physiopathologic conditions. We have recently demonstrated that IDO1 does not merely degrade tryptophan and produce immunoregulatory kynurenines, but it also acts as a signal-transducing molecule, independently of its enzymic function. IDO1 signalling activity is triggered in plasmacytoid dendritic cells (pDCs) by transforming growth factor-β (TGF-β), an event that requires the non-canonical NF-κB pathway and induces long-lasting IDO1 expression and autocrine TGF-β production in a positive feedback loop, thus sustaining a stably regulatory phenotype in pDCs. IDO1 expression and catalytic function are defective in pDCs from non-obese diabetic (NOD) mice, a prototypic model of autoimmune diabetes. In the present study, we found that TGF-β failed to activate IDO1 signalling function as well as up-regulate IDO1 expression in NOD pDCs. Moreover, TGF-β-treated pDCs failed to exert immunosuppressive properties in vivo. Nevertheless, transfection of NOD pDCs with Ido1 prior to TGF-β treatment resulted in activation of the Ido1 promoter and induction of non-canonical NF-κB and TGF-β, as well as decreased production of the pro-inflammatory cytokines, interleukin 6 (IL-6) and tumour necrosis factor-α (TNF-α). Overexpression of IDO1 in TGF-β-treated NOD pDCs also resulted in pDC ability to suppress the in vivo presentation of a pancreatic β-cell auto-antigen. Thus, our data suggest that a correction of IDO1 expression may restore its dual function and thus represent a proper therapeutic manoeuvre in this autoimmune setting.
Keywords: IDO1; autoimmune diabetes; immune regulation; non-canonical NF-κB; non-obese diabetic (NOD) mice; plasmacytoid dendritic cells; tryptophan catabolism.
© 2014 The Authors. Journal of Cellular and Molecular Medicine published by John Wiley & Sons Ltd and Foundation for Cellular and Molecular Medicine.
Figures
Similar articles
-
Chimeric Vaccine Stimulation of Human Dendritic Cell Indoleamine 2, 3-Dioxygenase Occurs via the Non-Canonical NF-κB Pathway.PLoS One. 2016 Feb 16;11(2):e0147509. doi: 10.1371/journal.pone.0147509. eCollection 2016. PLoS One. 2016. PMID: 26881431 Free PMC article.
-
Distinct roles of immunoreceptor tyrosine-based motifs in immunosuppressive indoleamine 2,3-dioxygenase 1.J Cell Mol Med. 2017 Jan;21(1):165-176. doi: 10.1111/jcmm.12954. Epub 2016 Sep 30. J Cell Mol Med. 2017. PMID: 27696702 Free PMC article.
-
Galectin 3 protects from cisplatin-induced acute kidney injury by promoting TLR-2-dependent activation of IDO1/Kynurenine pathway in renal DCs.Theranostics. 2019 Aug 14;9(20):5976-6001. doi: 10.7150/thno.33959. eCollection 2019. Theranostics. 2019. PMID: 31534532 Free PMC article.
-
Indoleamine 2,3-dioxygenase: from catalyst to signaling function.Eur J Immunol. 2012 Aug;42(8):1932-7. doi: 10.1002/eji.201242572. Eur J Immunol. 2012. PMID: 22865044 Review.
-
IDO and regulatory T cells: a role for reverse signalling and non-canonical NF-kappaB activation.Nat Rev Immunol. 2007 Oct;7(10):817-23. doi: 10.1038/nri2163. Nat Rev Immunol. 2007. PMID: 17767193 Review.
Cited by
-
Impact of IDO1 and IDO2 on the B Cell Immune Response.Front Immunol. 2022 Apr 13;13:886225. doi: 10.3389/fimmu.2022.886225. eCollection 2022. Front Immunol. 2022. PMID: 35493480 Free PMC article. Review.
-
The Coevolution of IDO1 and AhR in the Emergence of Regulatory T-Cells in Mammals.Front Immunol. 2015 Feb 12;6:58. doi: 10.3389/fimmu.2015.00058. eCollection 2015. Front Immunol. 2015. PMID: 25729384 Free PMC article. No abstract available.
-
Comparative genotypic and phenotypic analysis of human peripheral blood monocytes and surrogate monocyte-like cell lines commonly used in metabolic disease research.PLoS One. 2018 May 10;13(5):e0197177. doi: 10.1371/journal.pone.0197177. eCollection 2018. PLoS One. 2018. PMID: 29746559 Free PMC article.
-
Identification of a Compound Inhibiting Both the Enzymatic and Nonenzymatic Functions of Indoleamine 2,3-Dioxygenase 1.ACS Pharmacol Transl Sci. 2024 Sep 12;7(10):3056-3070. doi: 10.1021/acsptsci.4c00265. eCollection 2024 Oct 11. ACS Pharmacol Transl Sci. 2024. PMID: 39421661 Free PMC article.
-
A combination dual-sized microparticle system modulates dendritic cells and prevents type 1 diabetes in prediabetic NOD mice.Clin Immunol. 2015 Sep;160(1):90-102. doi: 10.1016/j.clim.2015.03.023. Epub 2015 Apr 2. Clin Immunol. 2015. PMID: 25842187 Free PMC article.
References
-
- Kishimoto H, Sprent J. A defect in central tolerance in NOD mice. Nat Immunol. 2001;2:1025–31. - PubMed
-
- Liu J, Beller D. Aberrant production of IL-12 by macrophages from several autoimmune-prone mouse strains is characterized by intrinsic and unique patterns of NF-κB expression and binding to the IL-12 p40 promoter. Immunol. 2002;169:581–6. - PubMed
-
- Suarez-Pinzon WL, Mabley JG, Strynadka K, et al. An inhibitor of inducible nitric oxide synthase and scavenger of peroxynitrite prevents diabetes development in NOD mice. J Autoimmun. 2001;16:449–55. - PubMed
-
- Serreze DV, Gaskins HR, Leiter EH. Defects in the differentiation and function of antigen presenting cells in NOD/Lt mice. J Immunol. 1993;150:2534–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
