

The intracellular Ca²⁺ channel MCOLN1 is required for sarcolemma repair to prevent muscular dystrophy
- PMID: 25216637
- PMCID: PMC4192061
- DOI: 10.1038/nm.3611
The intracellular Ca²⁺ channel MCOLN1 is required for sarcolemma repair to prevent muscular dystrophy
Abstract
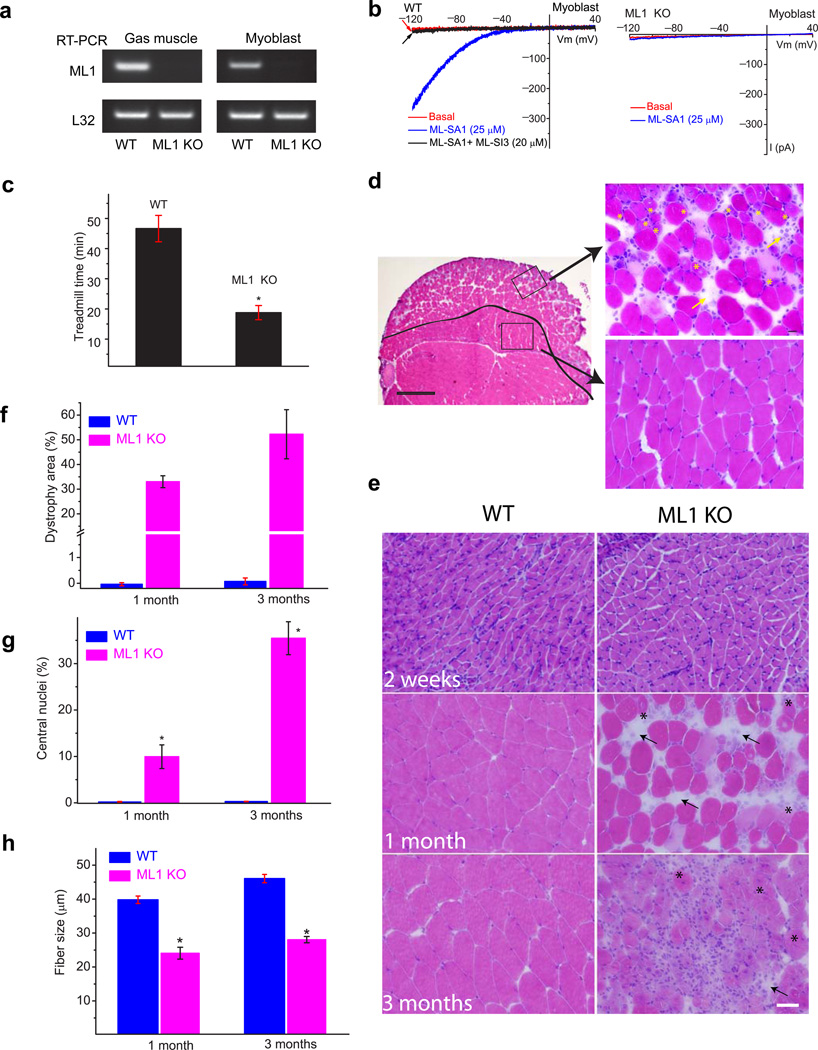
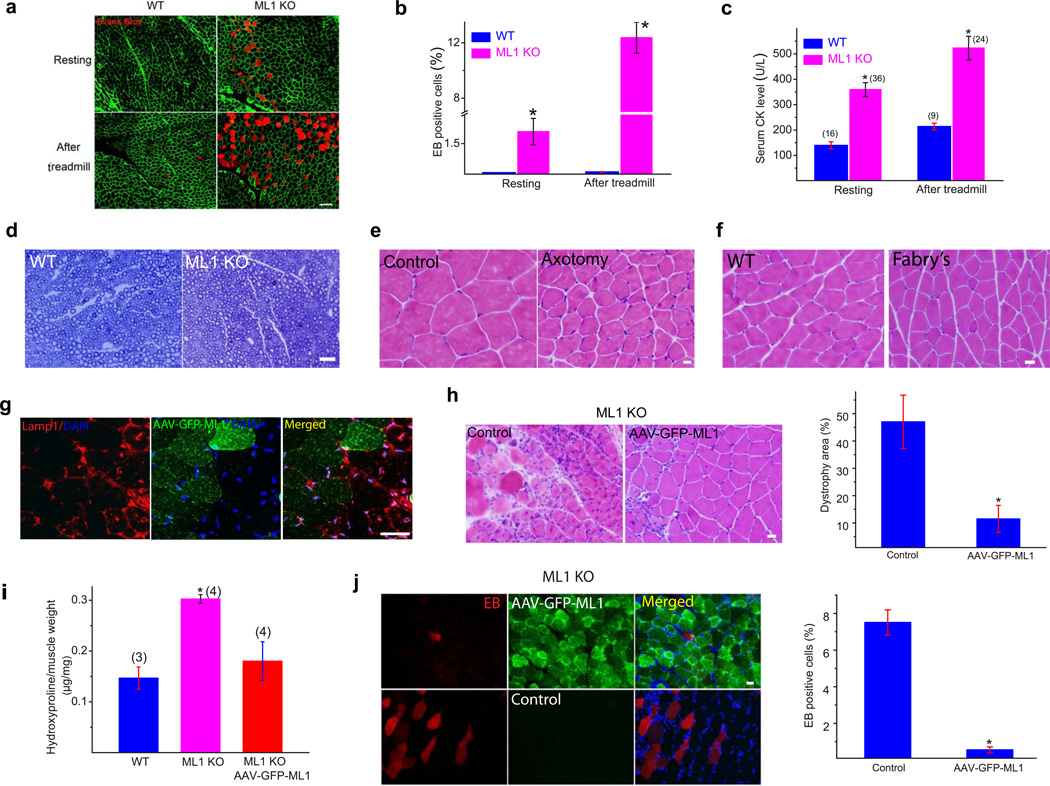
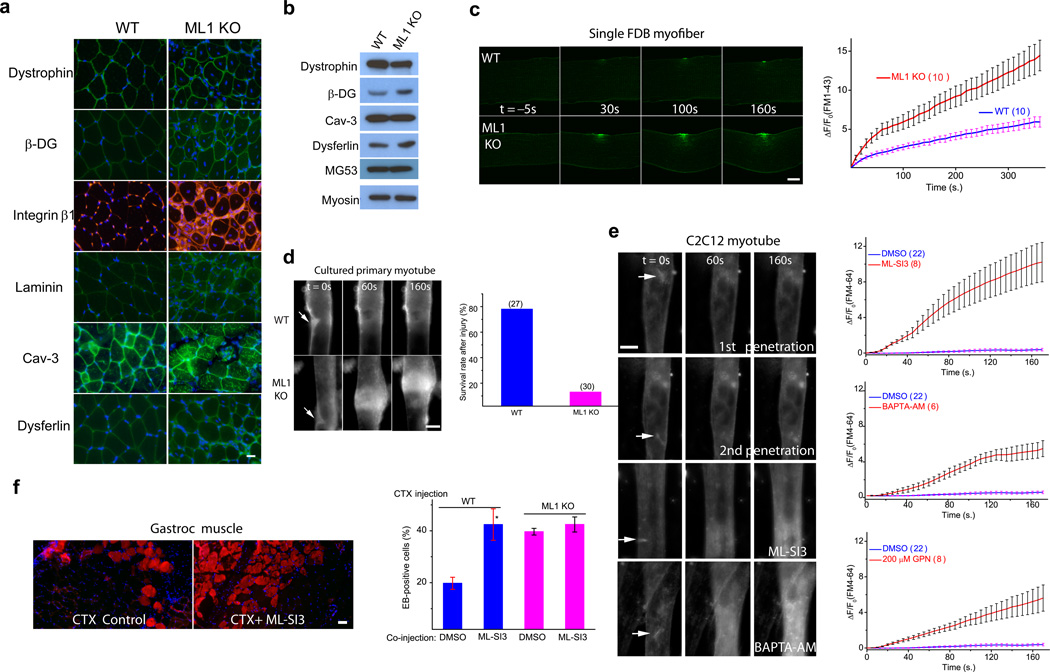
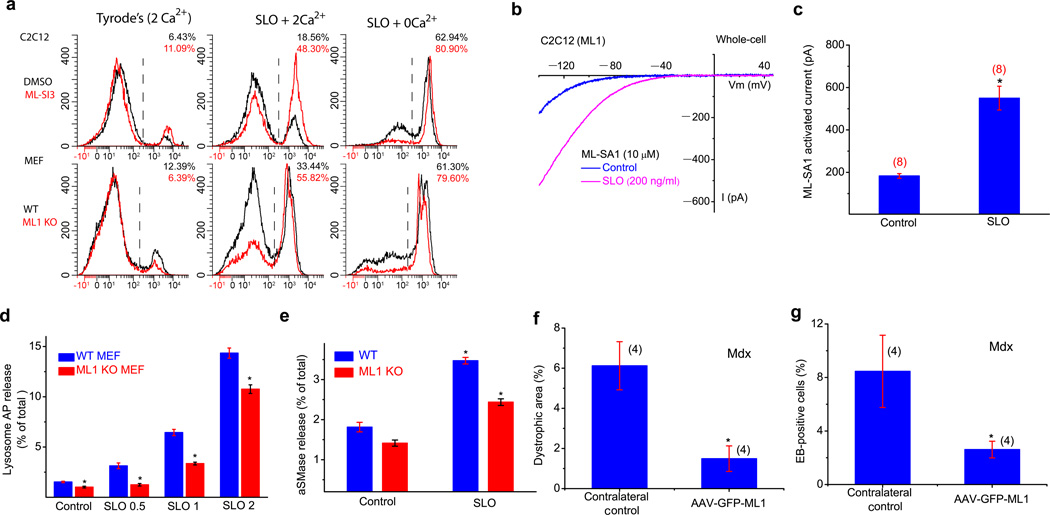
The integrity of the plasma membrane is maintained through an active repair process, especially in skeletal and cardiac muscle cells, in which contraction-induced mechanical damage frequently occurs in vivo. Muscular dystrophies (MDs) are a group of muscle diseases characterized by skeletal muscle wasting and weakness. An important cause of these group of diseases is defective repair of sarcolemmal injuries, which normally requires Ca(2+) sensor proteins and Ca(2+)-dependent delivery of intracellular vesicles to the sites of injury. MCOLN1 (also known as TRPML1, ML1) is an endosomal and lysosomal Ca(2+) channel whose human mutations cause mucolipidosis IV (ML4), a neurodegenerative disease with motor disabilities. Here we report that ML1-null mice develop a primary, early-onset MD independent of neural degeneration. Although the dystrophin-glycoprotein complex and the known membrane repair proteins are expressed normally, membrane resealing was defective in ML1-null muscle fibers and also upon acute and pharmacological inhibition of ML1 channel activity or vesicular Ca(2+) release. Injury facilitated the trafficking and exocytosis of vesicles by upmodulating ML1 channel activity. In the dystrophic mdx mouse model, overexpression of ML1 decreased muscle pathology. Collectively, our data have identified an intracellular Ca(2+) channel that regulates membrane repair in skeletal muscle via Ca(2+)-dependent vesicle exocytosis.
Figures
References
-
- Clarke MS, Khakee R, McNeil PL. Loss of cytoplasmic basic fibroblast growth factor from physiologically wounded myofibers of normal and dystrophic muscle. J Cell Sci. 1993;106(Pt 1):121–133. - PubMed
-
- Davies KE, Nowak KJ. Molecular mechanisms of muscular dystrophies: old and new players. Nat Rev Mol Cell Biol. 2006;7:762–773. - PubMed
-
- Bansal D, et al. Defective membrane repair in dysferlin-deficient muscular dystrophy. Nature. 2003;423:168–172. - PubMed
Method References
-
- Bolsover FE, Murphy E, Cipolotti L, Werring DJ, Lachmann RH. Cognitive dysfunction and depression in Fabry disease: a systematic review. J Inherit Metab Dis. 2013 - PubMed
-
- Springer ML, Rando TA, Blau HM. Gene delivery to muscle. Curr Protoc Hum Genet. 2002;Chapter 13(Unit13):14. - PubMed
-
- Brooke MH, Kaiser KK. Muscle fiber types: how many and what kind? Arch Neurol. 1970;23:369–379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
