

Shigella flexneri targets the HP1γ subcode through the phosphothreonine lyase OspF
- PMID: 25216677
- PMCID: PMC4282571
- DOI: 10.15252/embj.201489244
Shigella flexneri targets the HP1γ subcode through the phosphothreonine lyase OspF
Abstract
HP1 proteins are transcriptional regulators that, like histones, are targets for post-translational modifications defining an HP1-mediated subcode. HP1γ has multiple phosphorylation sites, including serine 83 (S83) that marks it to sites of active transcription. In a guinea pig model for Shigella enterocolitis, we observed that the defective type III secretion mxiD Shigella flexneri strain caused more HP1γ phosphorylation in the colon than the wild-type strain. Shigella interferes with HP1 phosphorylation by injecting the phospholyase OspF. This effector interacts with HP1γ and alters its phosphorylation at S83 by inactivating ERK and consequently MSK1, a downstream kinase. MSK1 that here arises as a novel HP1γ kinase, phosphorylates HP1γ at S83 in the context of an MSK1-HP1γ complex, and thereby favors its accumulation on its target genes. Genome-wide transcriptome analysis reveals that this mechanism is linked to up-regulation of proliferative gene and fine-tuning of immune gene expression. Thus, in addition to histones, bacteria control host transcription by modulating the activity of HP1 proteins, with potential implications in transcriptional reprogramming at the mucosal barrier.
Keywords: CBX3; Heterochromatin protein 1; MSK1; Shigella; colon.
© 2014 The Authors.
Figures
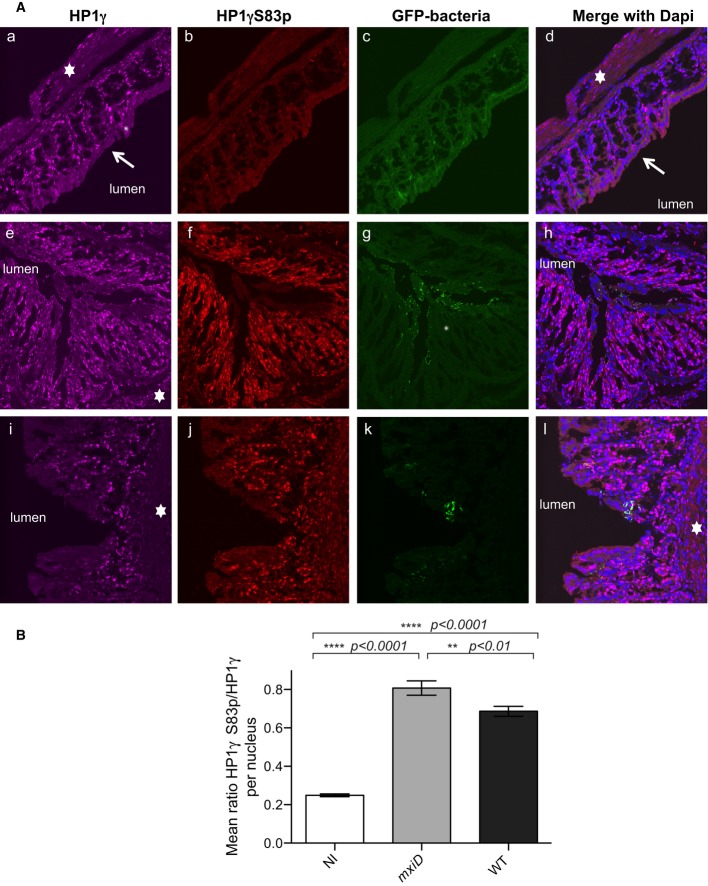
Samples of the distal colon were taken 7 h after infection with the T3SS defective mxiD Shigella pGFP(e–h), WT Shigella pGFP (i–l) or PBS treated as control (a–d) and co-stained with anti-HP1γ (a, e, and i) or anti-HP1γS83p (b, f, and j) antibodies and merged with DAPI (d, h, and l). GFP allowed for visualization of the bacterium (c, g, and k). The star indicated the submucosa, and the arrow showed the terminally differentiated columnar absorptive enterocytes devoid of HP1γ.
Fluorescence intensity ratios between HP1γS83p and total HP1γ. HP1γS83p staining was quantified on HP1γ-positive nuclei in each field. The intensity of fluorescence for each channel of interest was measured, and the ratio of fluorescence intensity between HP1γS83p and total HP1γ signals was calculated as described in Materials and Methods. Statistical analysis was performed as described in Materials and Methods.
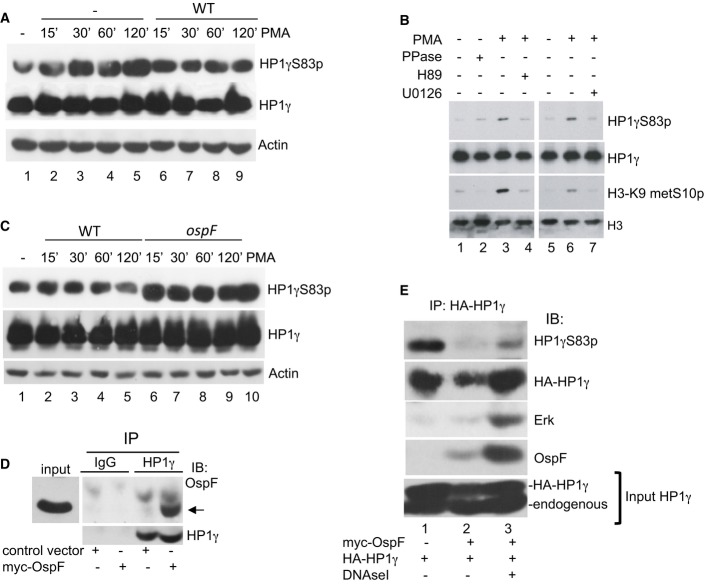
Shigella inhibits PMA-induced formation of HP1γS83p. HeLa cells were infected with the Shigella flexneri invasive strain (WT) and stimulated or not by PMA at the indicated times. Western blots were performed with anti-HP1γS83p, anti-HP1γ, and anti-actin antibodies.
Pharmacological inhibition of the MAPK/MSK1 pathway blocks formation of HP1γS83p. HeLa cells were pretreated for 1 h with the MEK1 inhibitor U0126 or MSK1 inhibitor H89 and stimulated by PMA. Western blots were performed with anti-HP1γS83p, anti-HP1γ, anti-H3K9me2S10p, or anti-H3 antibodies.
HeLa cells were infected with the Shigella flexneri invasive (WT) or the ospF-deficient (ospF) strains at the indicated time or stimulated by PMA (60 min). Western blots were performed with anti-HP1γS83p, anti-HP1γ, and anti-actin antibodies.
Immunoprecipitation of endogenous HP1γ from HeLa cells overexpressing OspF. HeLa cells were transiently transfected with the indicated plasmids. Cellular extract were immunoprecipitated with mouse IgG as a negative control or HP1γ antibodies. Western blots were performed with OspF and HP1γ antibodies. The arrow indicates the OspF signal.
Dephosphorylation at S83 in HP1γ immunoprecipitates containing OspF. HeLa cells were transiently transfected with the indicated plasmids and cellular extract immunoprecipitated with HP1γ antibody in the absence or the presence of DNase I. Western blots were performed with anti-HP1γS83p, anti-HP1γ, anti-OspF, or anti-ERK antibodies.
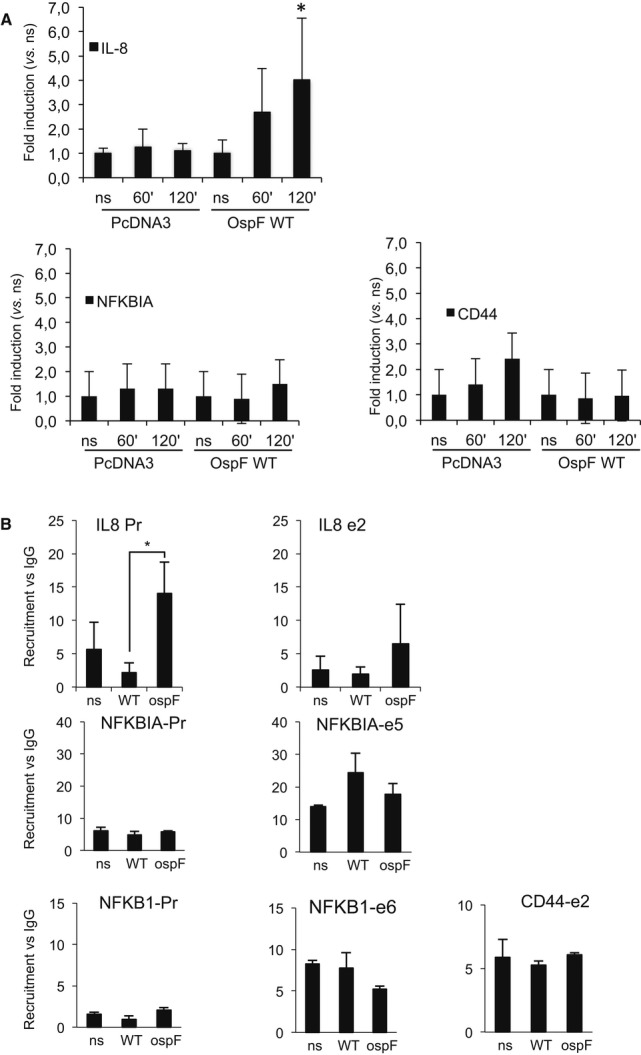
HeLa cells were transfected with the indicated plasmids and stimulated by TNF at the indicated times. Chromatin immunoprecipitation experiments (ChIP) were performed using anti-OspF antibodies. Enrichment in chromatin was quantified by qPCR using indicated primers at the IL8, CD44, and NFKBIA genes. Enrichment values are the mean of three independent experiments. *P < 0.05 when compared to the non-stimulated cells (ns).
OspF modulates HP1γ chromatin association. HeLa cells were infected with the Shigella flexneri invasive strain (WT) or the OspF-deficient strain (ospF). ChiPs were performed using anti-HP1γ antibodies or mouse IgG as control. Enrichment in chromatin was quantified by qPCR using indicated primers at the IL8, NFKB1, and NFKBIA genes (Pr, proximal promoter; e2, exon 2). Enrichment values are the mean of three independent experiments. Significance of the differences was estimated using Student's t-test with a threshold at 5%.
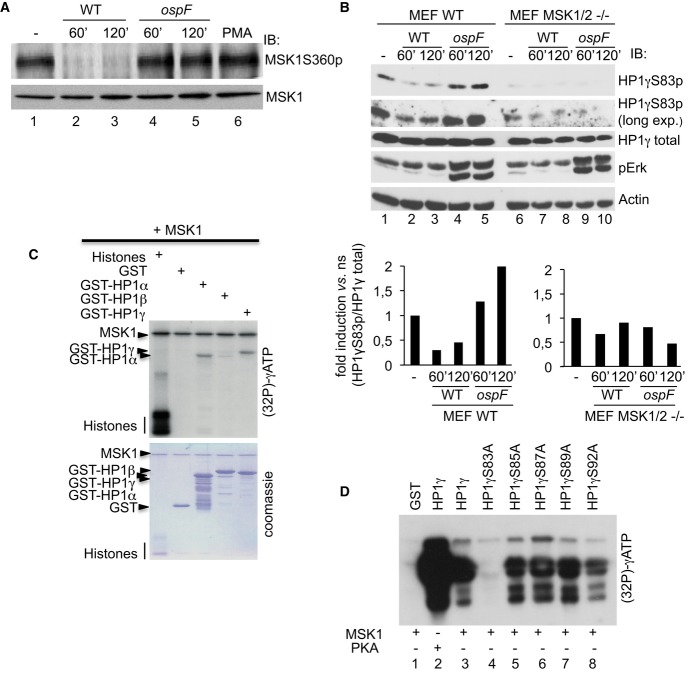
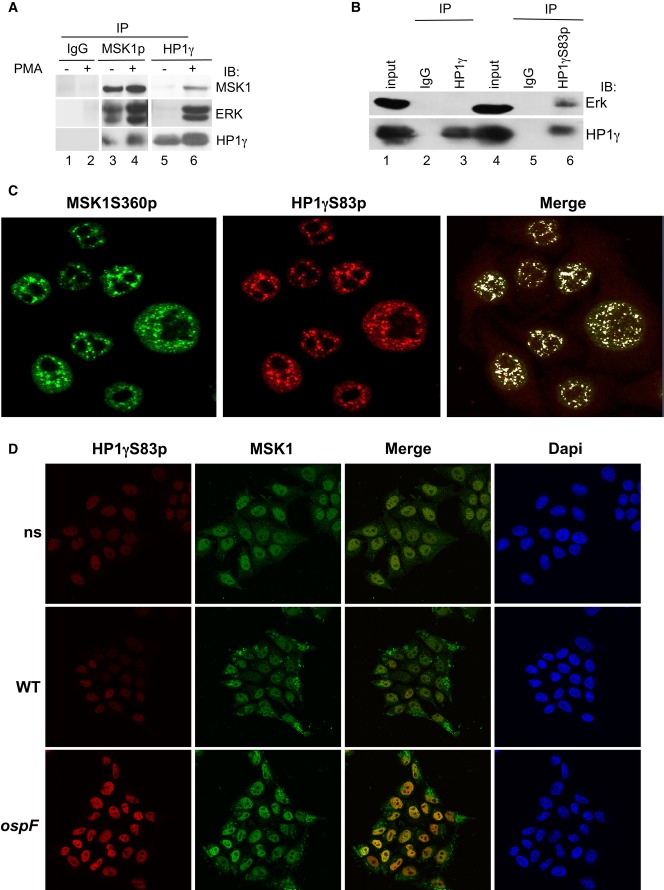
PMA stimulation leads to the formation of molecular tri-complex between the HP1γ and the ERK/MSK1 kinases. HeLa cells were stimulation by PMA (60 min), and cellular extracts were immunoprecipitated with mouse IgG as a negative control or anti-MSK1S360p or anti-HP1γ antibodies. Western blots were performed with anti-MSK1, anti-ERK, and anti-HP1γ antibodies.
The phosphorylated pool of HP1γ pulls down the ERK kinase. Cellular extracts from HeLa cells were immunoprecipitated with mouse IgG as a negative control or anti-HP1γS83p, or anti-HP1γ antibodies. Western blots were performed with anti-ERK and anti-HP1γS83p antibodies.
Confocal microscopy showing high coincidence of immunostaining between HP1γS83p and active MSK1. Immunofluorescence was performed with anti-HP1γS83p (red) and anti-MSK1S360p (green) antibodies.
Confocal microscopy showing the immunostaining between HP1γS83p (red) and MSK1 upon infection with the WT and ospF mutant strains (green).
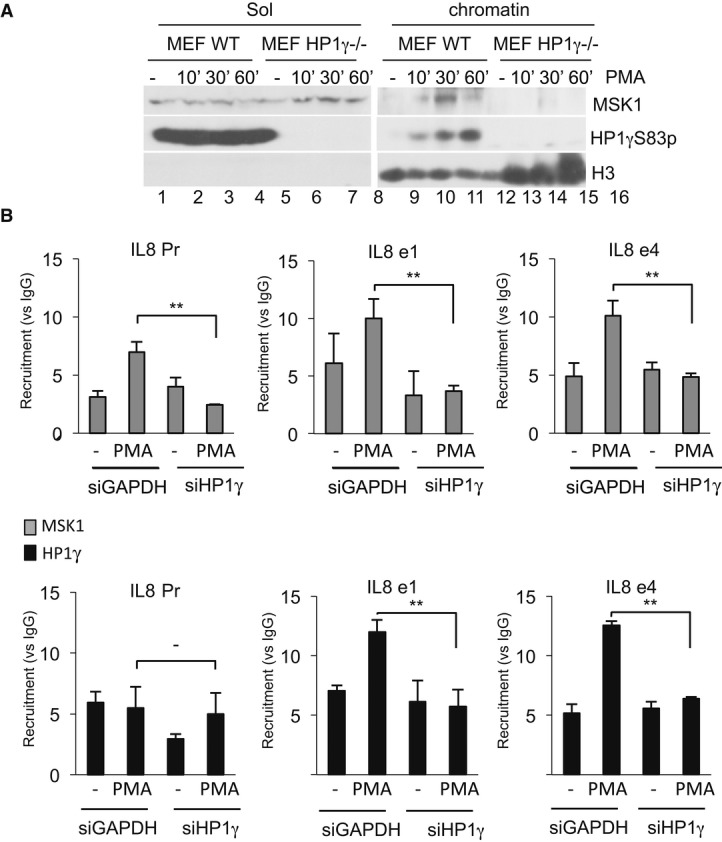
WT and HP1γ null mouse fibroblasts were stimulated by PMA at the indicated times. Nuclear extract were processed as indicated in Materials and Methods. Western blots were performed with anti-MSK1, anti-HP1γS83p, and anti-histone H3 antibodies.
HeLa cells were transfected with siHP1γ or, as a control, siGAPDH and stimulated by PMA (40 min). ChIPs were performed using anti-HP1γ or anti-MSK1 antibodies, or unrelated mouse IgGs as a control. Results are expressed as fold induction versus IgG for each point (**P < 0.01). Enrichment values are the mean of two independent experiments. Significance of the differences was estimated using Student's t-test with a threshold at 5%.
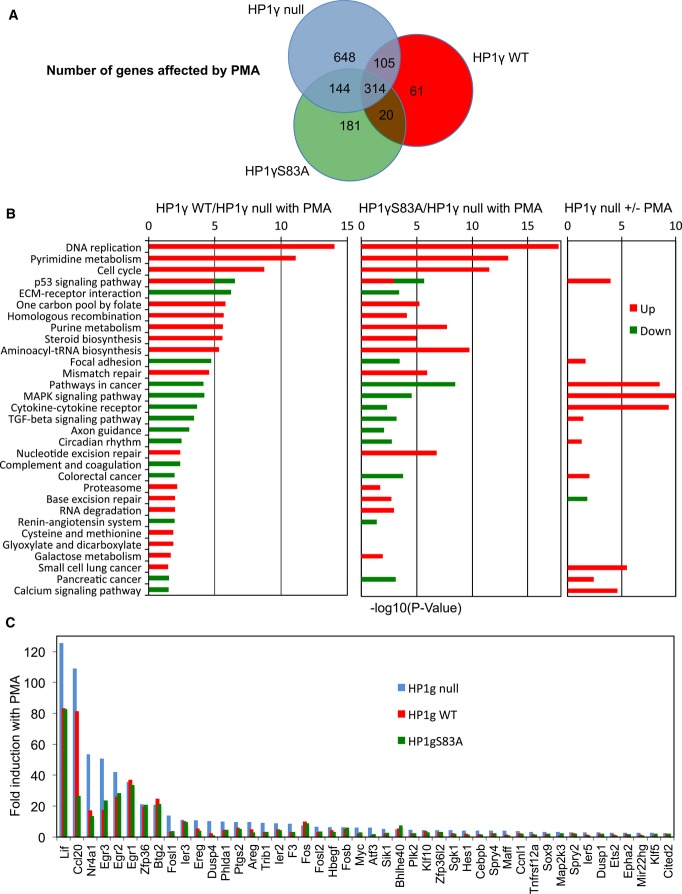
Venn diagram showing the number of transcripts regulated by PMA stimulation in HP1γ null (blue), HP1γ WT (red), and HP1γS83A (green) MEF-derived cell lines (≥ 1.5-fold change in expression, ≥ 20 reads, P < 0.05).
KEGG functional categories significantly enriched (P < 0.05) for genes differentially expressed in the indicated conditions, depicted according to their P-value [−log10(P-value)].
Fold induction by PMA of a subset of OspF target genes (≥ 2-fold change in expression in HP1γ null) in each of the indicated cell lines. Data and P-values are provided in Supplementary Table S1.
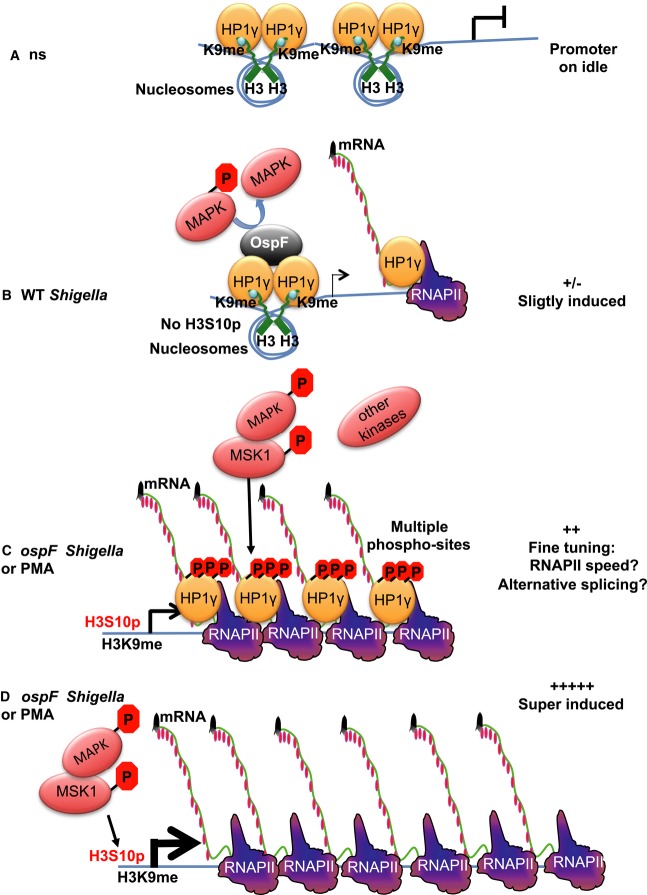
In unstimulated cells (ns), HP1 is brought to the promoter by histone H3K9 methylation and acts as a transcriptional repressor.
Shigella flexneri infection results in activation of the MAPK/MSK1 pathway, but this activation is rapidly dampened by OspF that dephosphorylates ERK. The combined effect of these two events is a moderate transcriptional activation of IL8. This causes HP1γ to leave the promoter, while inactivation of ERK prevents it from getting phosphorylated and join the transcription machinery. Consequently, accumulation of HP1γ at the IL8 gene drops.
Upon ospF Shigella infection or treatment with PMA, the MAPK nuclear pathway is fully triggered, levels of histone H3 marks permissive for transcription are increased (H3S10), and the transcription machinery is recruited. Activation of the MAPK nuclear pathway induces HP1γ phosphorylation and allows for its recruitment to IL8 and other similarly regulated immune genes.
In MEF KO cells, the loss of HP1γ eliminates an important checkpoint in the control of immune gene activation, leading to a deregulated possibly excessive gene expression in response to MAPK activation.
Comment in
-
Shigella hacks host immune responses by reprogramming the host epigenome.EMBO J. 2014 Nov 18;33(22):2598-600. doi: 10.15252/embj.201489934. Epub 2014 Sep 22. EMBO J. 2014. PMID: 25246515 Free PMC article.
-
Targeting of chromatin readers: a novel strategy used by the Shigella flexneri virulence effector OspF to reprogram transcription.Microb Cell. 2014 Dec 28;2(1):26-28. doi: 10.15698/mic2015.01.183. Microb Cell. 2014. PMID: 28357260 Free PMC article.
References
-
- Abe K, Naruse C, Kato T, Nishiuchi T, Saitou M, Asano M. Loss of heterochromatin protein 1 gamma reduces the number of primordial germ cells via impaired cell cycle progression in mice. Biol Reprod. 2011;85:1013–1024. - PubMed
-
- Allaoui A, Sansonetti PJ, Parsot C. MxiD, an outer membrane protein necessary for the secretion of the Shigella flexneri lpa invasins. Mol Microbiol. 1993;7:59–68. - PubMed
-
- Ameyar-Zazoua M, Rachez C, Souidi M, Robin P, Fritsch L, Young R, Morozova N, Fenouil R, Descostes N, Andrau JC, Mathieu J, Hamiche A, Ait-Si-Ali S, Muchardt C, Batsché E, Harel-Bellan A. Argonaute proteins couple chromatin silencing to alternative splicing. Nat Struct Mol Biol. 2012;19:998–1004. - PubMed
-
- Ananieva O, Darragh J, Johansen C, Carr JM, McIlrath J, Park JM, Wingate A, Monk CE, Toth R, Santos SG, Iversen L, Arthur JS. The kinases MSK1 and MSK2 act as negative regulators of Toll-like receptor signaling. Nat Immunol. 2008;9:1028–1036. - PubMed
-
- Arbibe L, Kim DW, Batsche E, Pedron T, Mateescu B, Muchardt C, Parsot C, Sansonetti PJ. An injected bacterial effector targets chromatin access for transcription factor NF-kappaB to alter transcription of host genes involved in immune responses. Nat Immunol. 2007;8:47–56. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
