

c-Jun activation in Schwann cells protects against loss of sensory axons in inherited neuropathy
- PMID: 25216747
- PMCID: PMC4208468
- DOI: 10.1093/brain/awu257
c-Jun activation in Schwann cells protects against loss of sensory axons in inherited neuropathy
Abstract
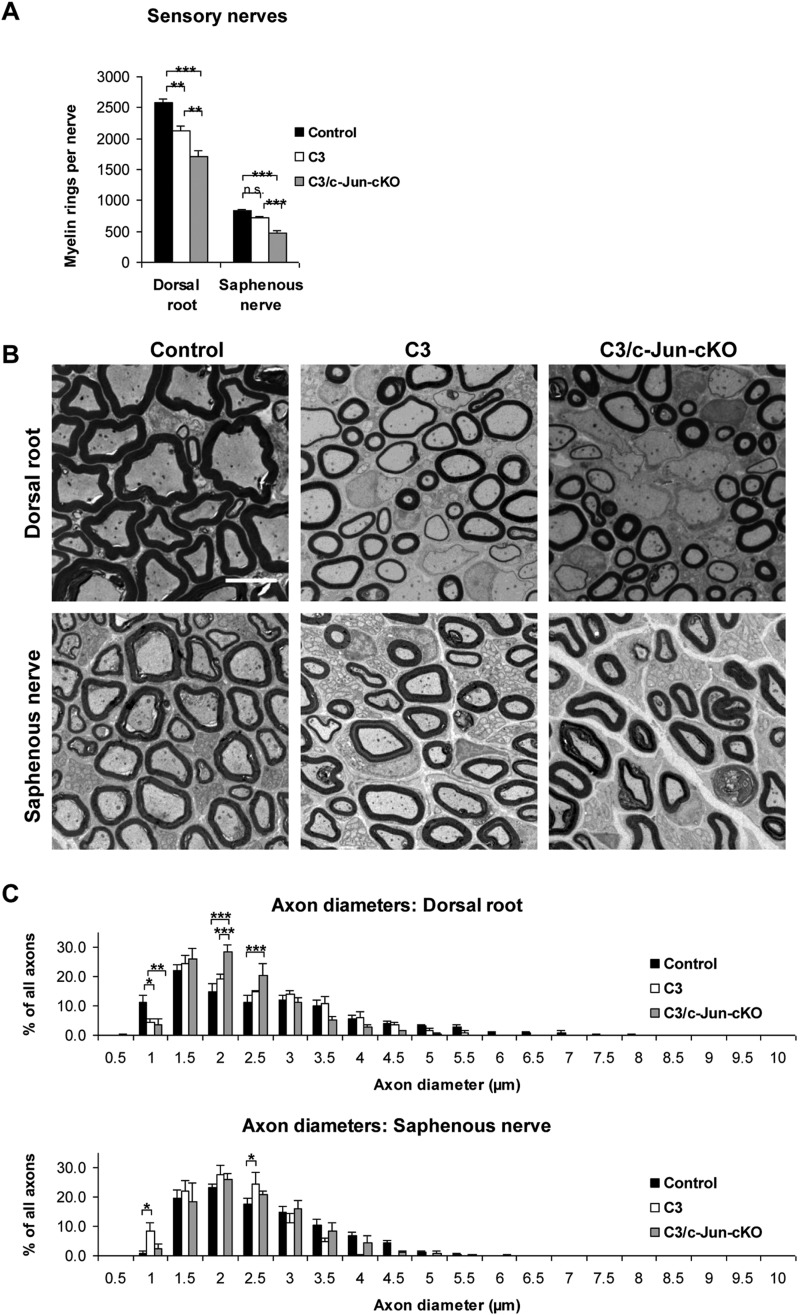
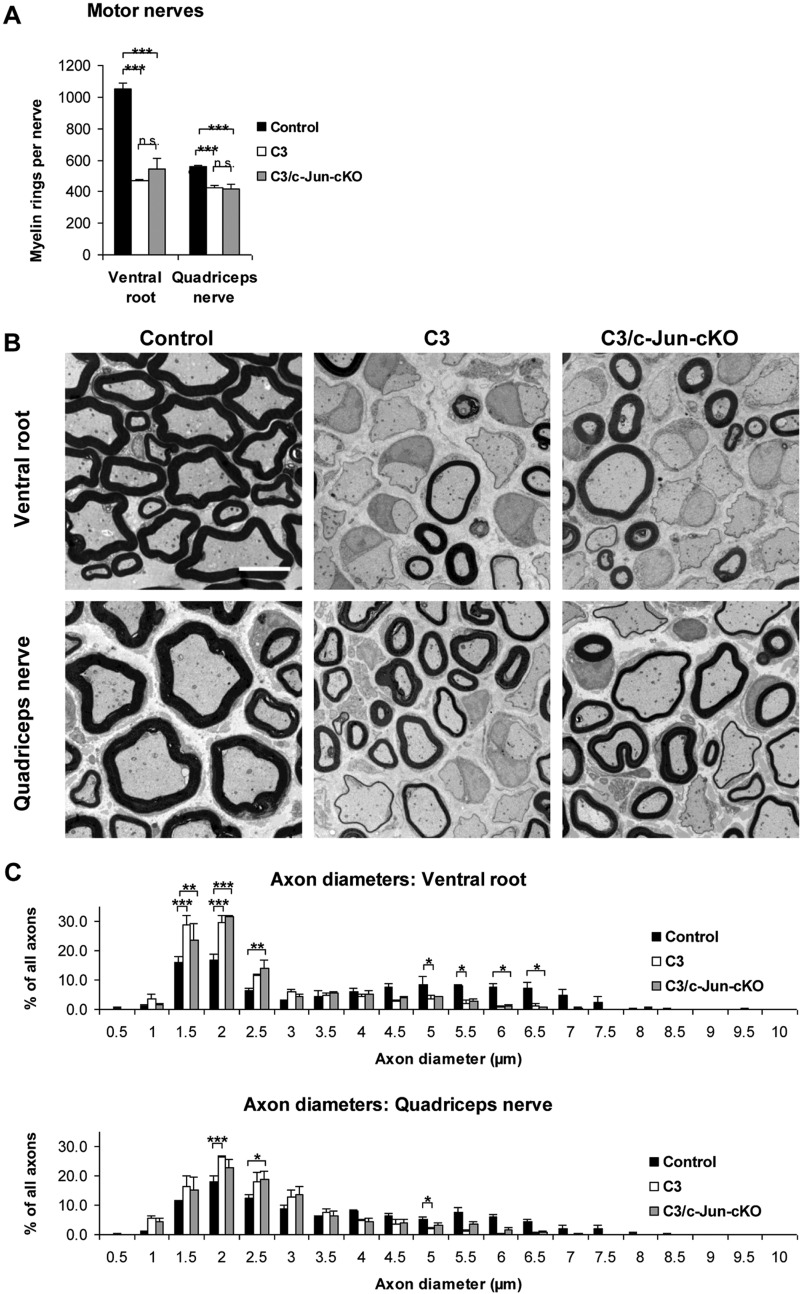
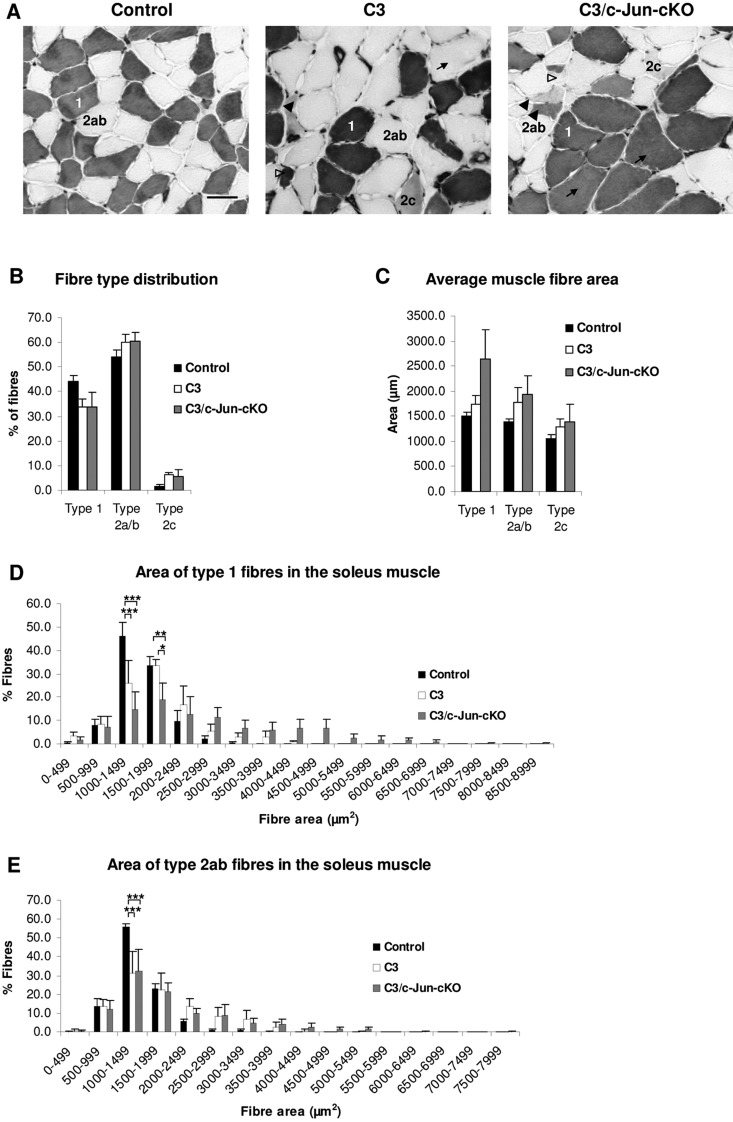
Charcot-Marie-Tooth disease type 1A is the most frequent inherited peripheral neuropathy. It is generally due to heterozygous inheritance of a partial chromosomal duplication resulting in over-expression of PMP22. A key feature of Charcot-Marie-Tooth disease type 1A is secondary death of axons. Prevention of axonal loss is therefore an important target of clinical intervention. We have previously identified a signalling mechanism that promotes axon survival and prevents neuron death in mechanically injured peripheral nerves. This work suggested that Schwann cells respond to injury by activating/enhancing trophic support for axons through a mechanism that depends on upregulation of the transcription factor c-Jun in Schwann cells, resulting in the sparing of axons that would otherwise die. As c-Jun orchestrates Schwann cell support for distressed neurons after mechanical injury, we have now asked: do Schwann cells also activate a c-Jun dependent neuron-supportive programme in inherited demyelinating disease? We tested this by using the C3 mouse model of Charcot-Marie-Tooth disease type 1A. In line with our previous findings in humans with Charcot-Marie-Tooth disease type 1A, we found that Schwann cell c-Jun was elevated in (uninjured) nerves of C3 mice. We determined the impact of this c-Jun activation by comparing C3 mice with double mutant mice, namely C3 mice in which c-Jun had been conditionally inactivated in Schwann cells (C3/Schwann cell-c-Jun(-/-) mice), using sensory-motor tests and electrophysiological measurements, and by counting axons in proximal and distal nerves. The results indicate that c-Jun elevation in the Schwann cells of C3 nerves serves to prevent loss of myelinated sensory axons, particularly in distal nerves, improve behavioural symptoms, and preserve F-wave persistence. This suggests that Schwann cells have two contrasting functions in Charcot-Marie-Tooth disease type 1A: on the one hand they are the genetic source of the disease, on the other, they respond to it by mounting a c-Jun-dependent response that significantly reduces its impact. Because axonal death is a central feature of much nerve pathology it will be important to establish whether an axon-supportive Schwann cell response also takes place in other conditions. Amplification of this axon-supportive mechanism constitutes a novel target for clinical intervention that might be useful in Charcot-Marie-Tooth disease type 1A and other neuropathies that involve axon loss.
Keywords: axonal degeneration; demyelinating disease; neural repair; neuron-glial interaction; neuropathy.
© The Author (2014). Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
Figures
References
-
- Birouk N, Gouider R, Le Guern E, Gugenheim M, Tardieu S, Maisonobe T, et al. Charcot-Marie-Tooth disease type 1A with 17p11.2 duplication. Clinical and electrophysiological phenotype study and factors influencing disease severity in 119 cases. Brain. 1997;120:813–23. - PubMed
-
- Brooke MH, Kaiser KK. Muscle fiber types: how many and what kind? Arch Neurol. 1970;23:369–79. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
