

Adjustment of host cells for accommodation of symbiotic bacteria: vacuole defunctionalization, HOPS suppression, and TIP1g retargeting in Medicago
- PMID: 25217511
- PMCID: PMC4213156
- DOI: 10.1105/tpc.114.128736
Adjustment of host cells for accommodation of symbiotic bacteria: vacuole defunctionalization, HOPS suppression, and TIP1g retargeting in Medicago
Abstract
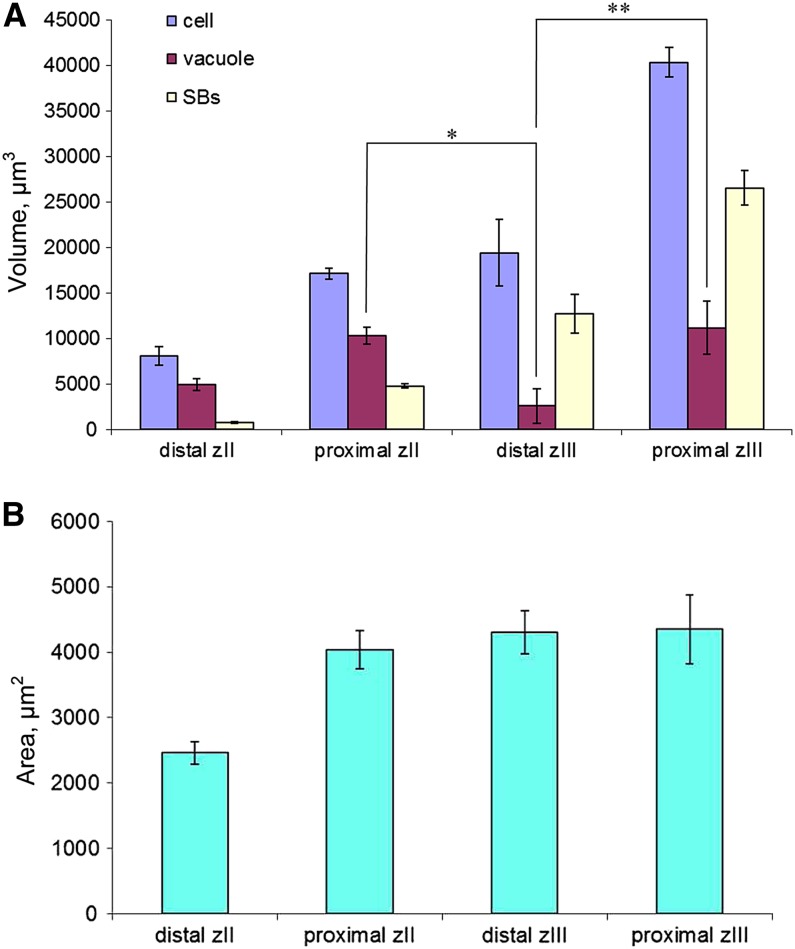
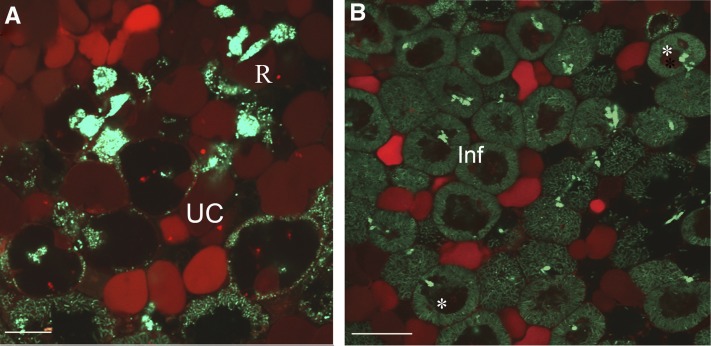
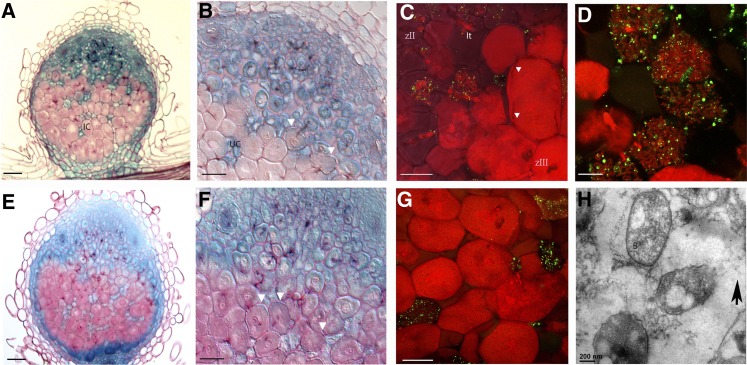
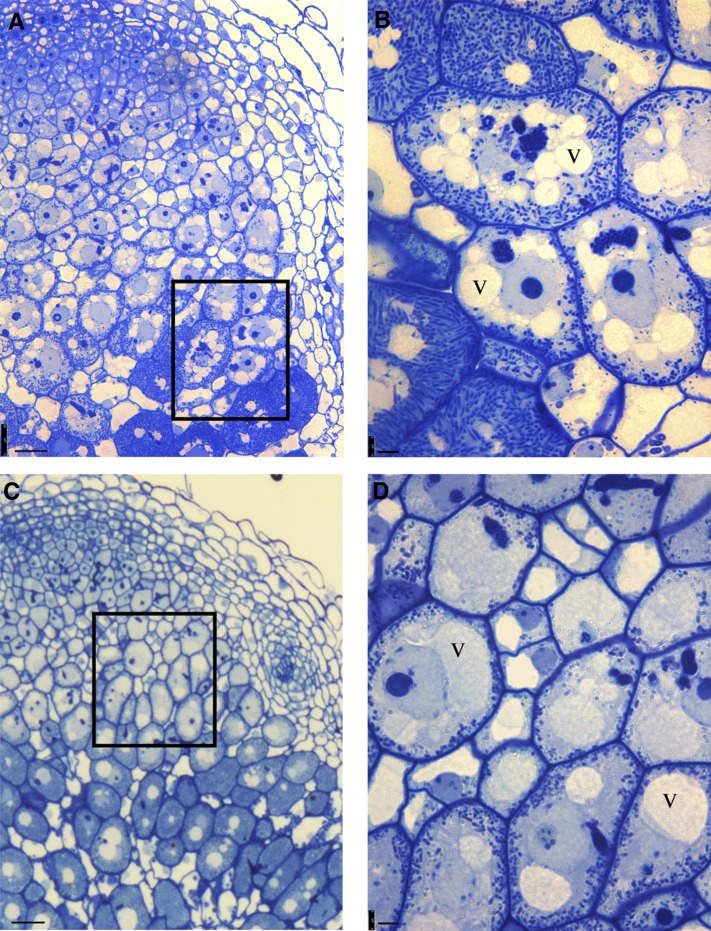
In legume-rhizobia symbioses, the bacteria in infected cells are enclosed in a plant membrane, forming organelle-like compartments called symbiosomes. Symbiosomes remain as individual units and avoid fusion with lytic vacuoles of host cells. We observed changes in the vacuole volume of infected cells and thus hypothesized that microsymbionts may cause modifications in vacuole formation or function. To examine this, we quantified the volumes and surface areas of plant cells, vacuoles, and symbiosomes in root nodules of Medicago truncatula and analyzed the expression and localization of VPS11 and VPS39, members of the HOPS vacuole-tethering complex. During the maturation of symbiosomes to become N2-fixing organelles, a developmental switch occurs and changes in vacuole features are induced. For example, we found that expression of VPS11 and VPS39 in infected cells is suppressed and host cell vacuoles contract, permitting the expansion of symbiosomes. Trafficking of tonoplast-targeted proteins in infected symbiotic cells is also altered, as shown by retargeting of the aquaporin TIP1g from the tonoplast membrane to the symbiosome membrane. This retargeting appears to be essential for the maturation of symbiosomes. We propose that these alterations in the function of the vacuole are key events in the adaptation of the plant cell to host intracellular symbiotic bacteria.
© 2014 American Society of Plant Biologists. All rights reserved.
Figures

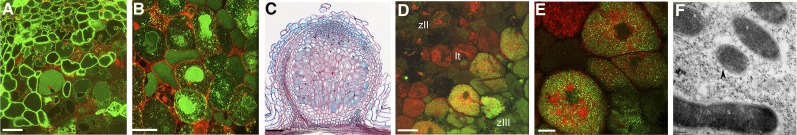
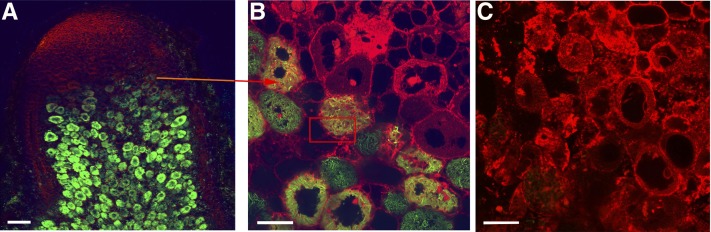
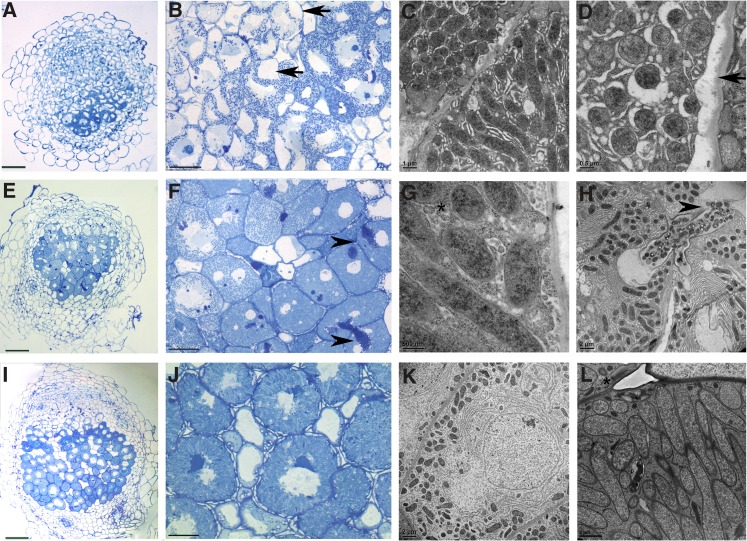
References
-
- Avila-Ospina, L., Moison, M., Yoshimoto, K., Masclaux-Daubresse, C. (2014). Autophagy, plant senescence, and nutrient recycling. J. Exp. Bot. 65: 3799–3811 - PubMed
-
- Balderhaar, H.J., Ungermann, C. (2013). CORVET and HOPS tethering complexes—Coordinators of endosome and lysosome fusion. J. Cell Sci. 126: 1307–1316 - PubMed
-
- Bassham, D.C., Laporte, M., Marty, F., Moriyasu, Y., Ohsumi, Y., Olsen, L.J., Yoshimoto, K. (2006). Autophagy in development and stress responses of plants. Autophagy 2: 2–11 - PubMed
-
- Behnia, R., Munro, S. (2005). Organelle identity and the signposts for membrane traffic. Nature 438: 597–604 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
