

Serine integrase chimeras with activity in E. coli and HeLa cells
- PMID: 25217617
- PMCID: PMC4197438
- DOI: 10.1242/bio.20148748
Serine integrase chimeras with activity in E. coli and HeLa cells
Abstract
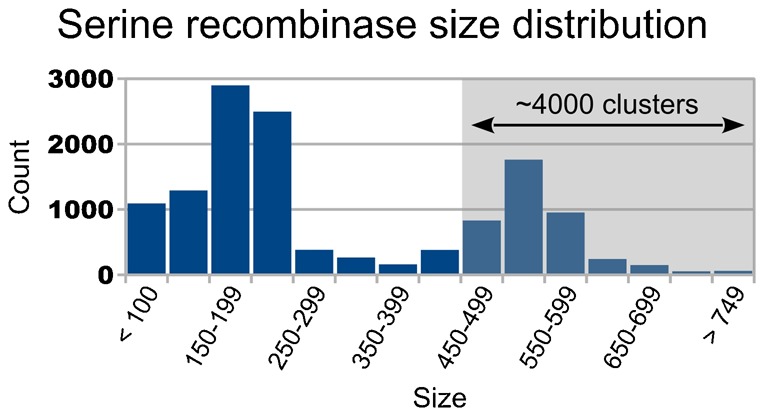
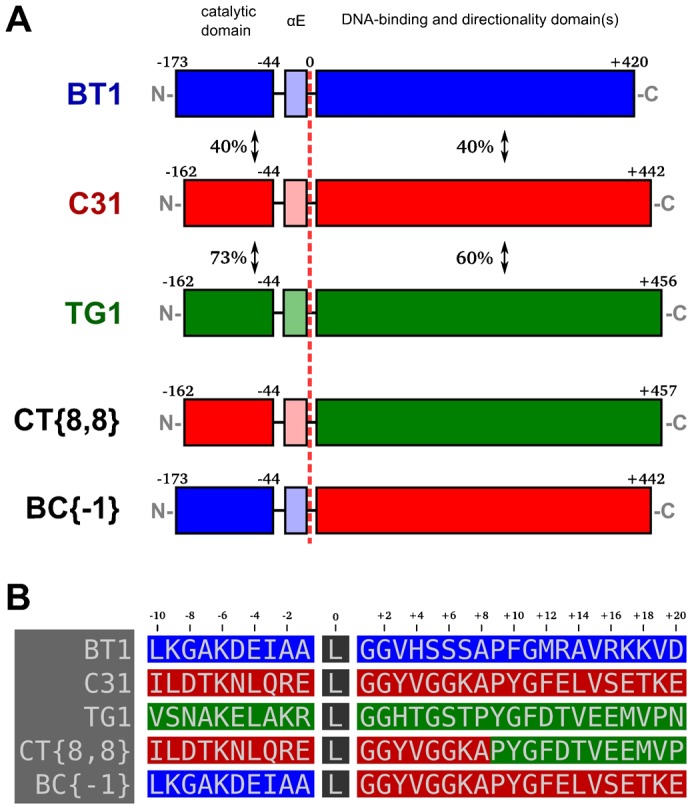
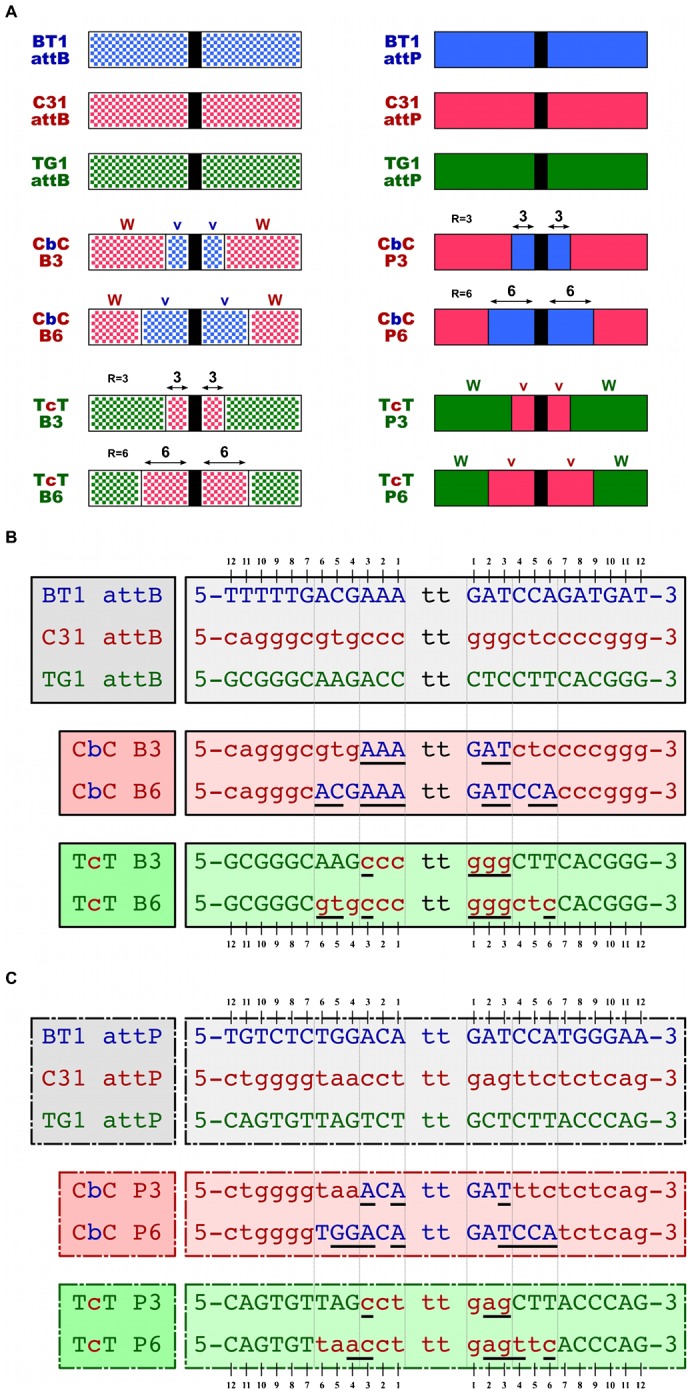
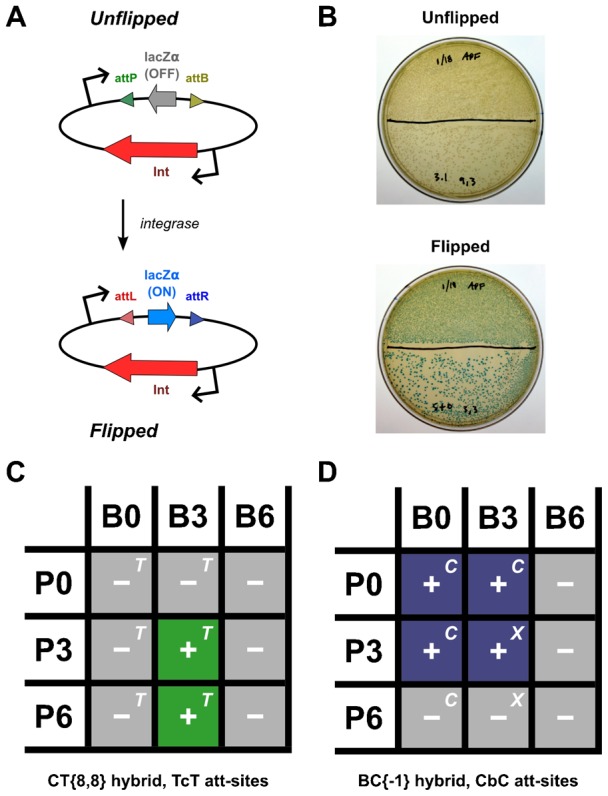
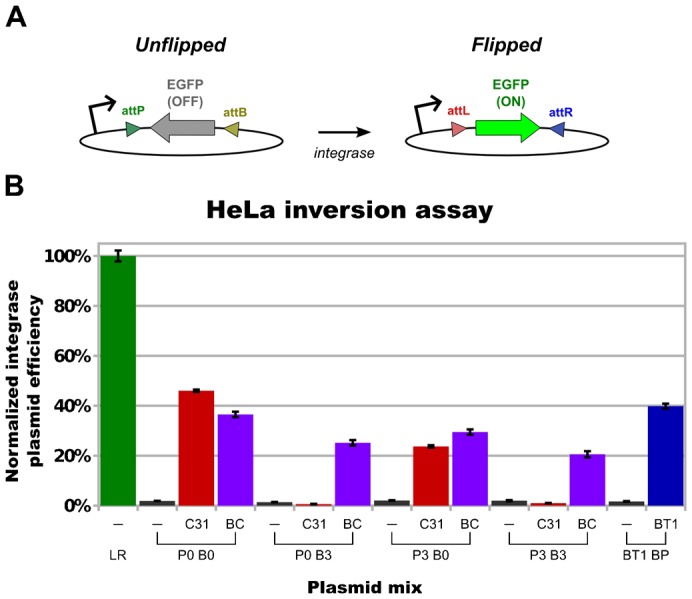
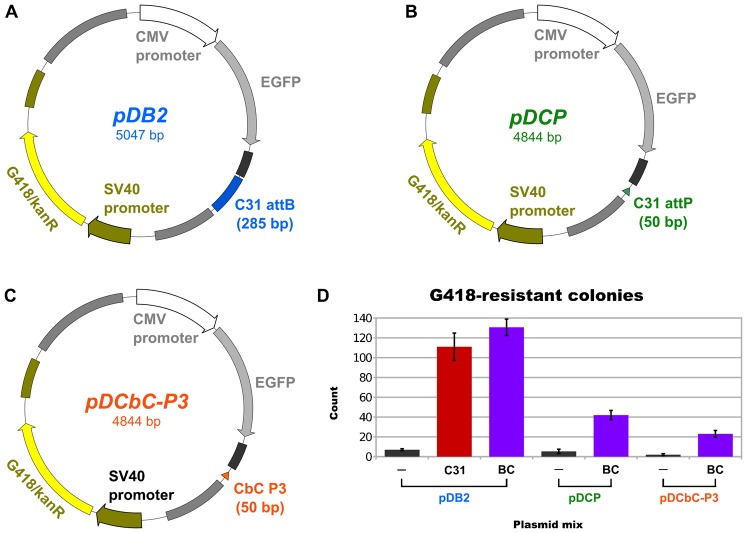
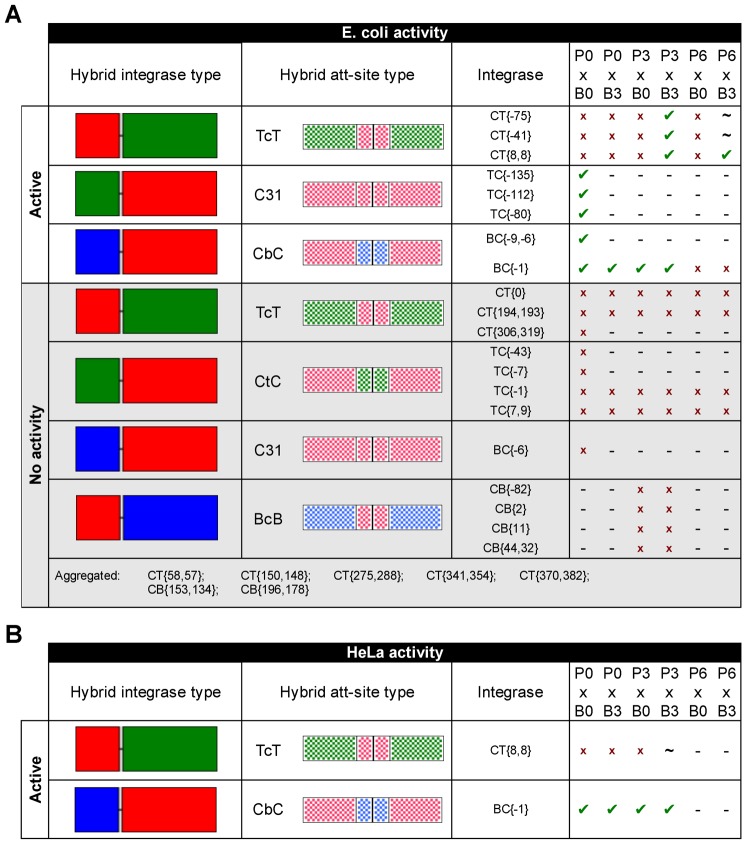
In recent years, application of serine integrases for genomic engineering has increased in popularity. The factor-independence and unidirectionality of these large serine recombinases makes them well suited for reactions such as site-directed vector integration and cassette exchange in a wide variety of organisms. In order to generate information that might be useful for altering the specificity of serine integrases and to improve their efficiency, we tested a hybridization strategy that has been successful with several small serine recombinases. We created chimeras derived from three characterized members of the serine integrase family, phiC31, phiBT1, and TG1 integrases, by joining their amino- and carboxy-terminal portions. We found that several phiBT1-phiC31 (BC) and phiC31-TG1 (CT) hybrid integrases are active in E. coli. BC chimeras function on native att-sites and on att-sites that are hybrids between those of the two donor enzymes, while CT chimeras only act on the latter att-sites. A BC hybrid, BC{-1}, was also active in human HeLa cells. Our work is the first to demonstrate chimeric serine integrase activity. This analysis sheds light on integrase structure and function, and establishes a potentially tractable means to probe the specificity of the thousands of putative large serine recombinases that have been revealed by bioinformatics studies.
Keywords: Chimeric recombinase; Genome engineering; Hybrid protein; Phage integrase; Sequence-specific recombination.
© 2014. Published by The Company of Biologists Ltd.
Conflict of interest statement
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
