

Notch directly regulates the cell morphogenesis genes Reck, talin and trio in adult muscle progenitors
- PMID: 25217625
- PMCID: PMC4215712
- DOI: 10.1242/jcs.151787
Notch directly regulates the cell morphogenesis genes Reck, talin and trio in adult muscle progenitors
Abstract
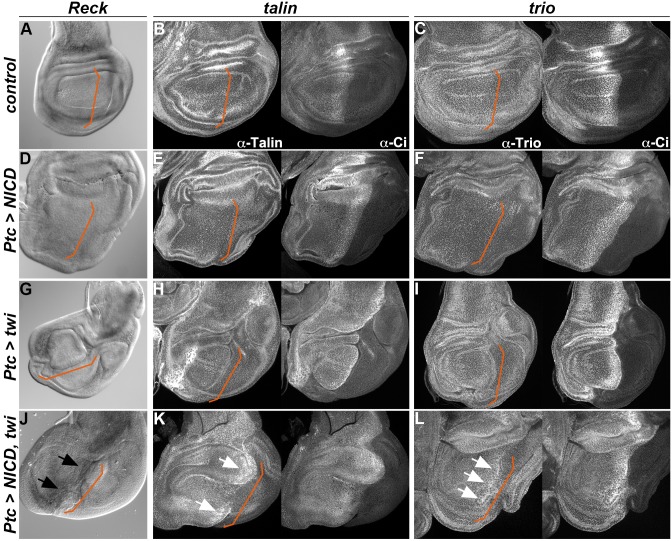
There is growing evidence that activation of the Notch pathway can result in consequences on cell morphogenesis and behaviour, both during embryonic development and cancer progression. In general, Notch is proposed to coordinate these processes by regulating expression of key transcription factors. However, many Notch-regulated genes identified in genome-wide studies are involved in fundamental aspects of cell behaviour, suggesting a more direct influence on cellular properties. By testing the functions of 25 such genes we confirmed that 12 are required in developing adult muscles, consistent with roles downstream of Notch. Focusing on three, Reck, rhea/talin and trio, we verify their expression in adult muscle progenitors and identify Notch-regulated enhancers in each. Full activity of these enhancers requires functional binding sites for Su(H), the DNA-binding transcription factor in the Notch pathway, validating their direct regulation. Thus, besides its well-known roles in regulating the expression of cell-fate-determining transcription factors, Notch signalling also has the potential to directly affect cell morphology and behaviour by modulating expression of genes such as Reck, rhea/talin and trio. This sheds new light on the functional outputs of Notch activation in morphogenetic processes.
Keywords: Drosophila; Gene regulation; Myogenesis; Notch; Reck; Talin; Trio.
© 2014. Published by The Company of Biologists Ltd.
Figures
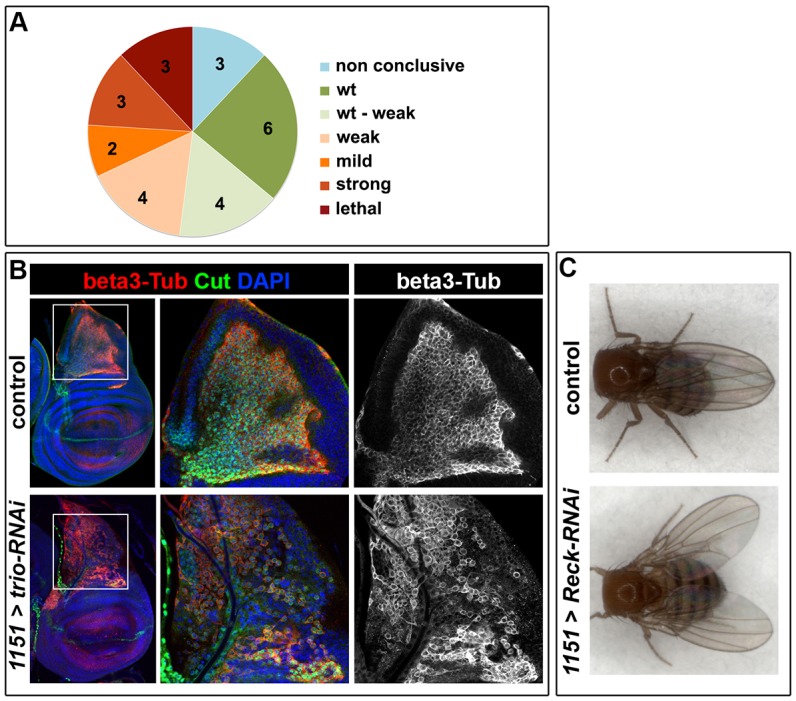
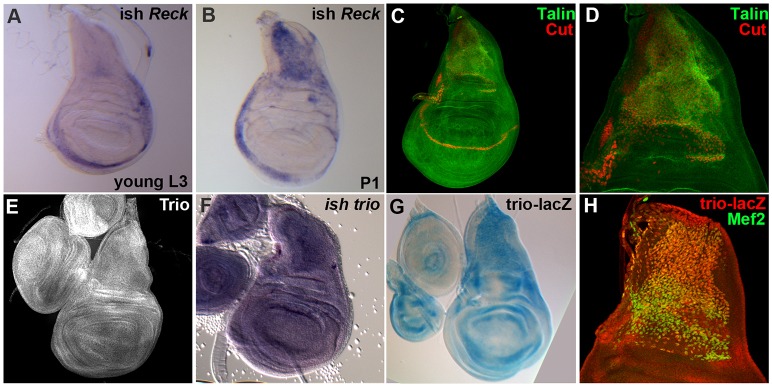
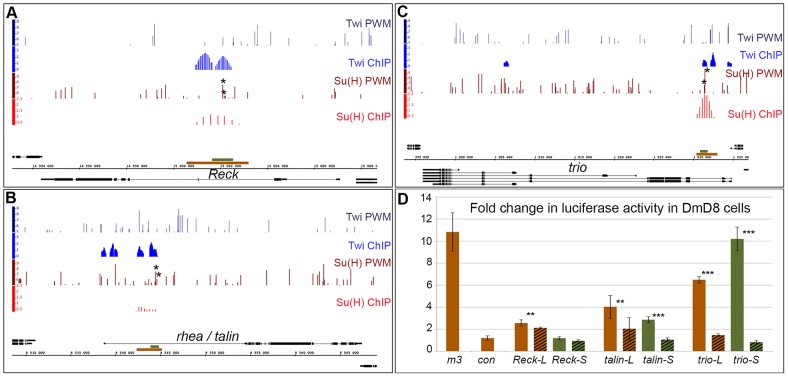
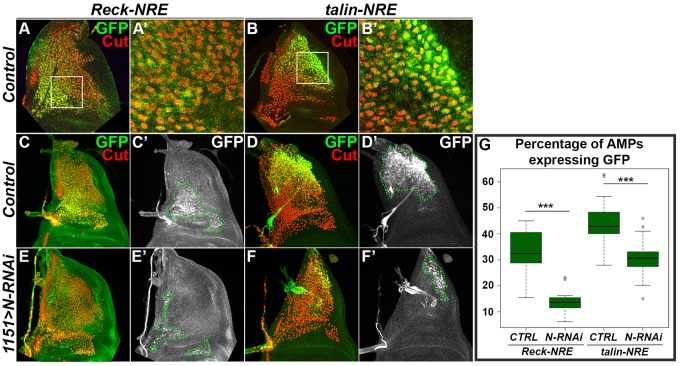
References
-
- Anant S., Roy S., VijayRaghavan K. (1998). Twist and Notch negatively regulate adult muscle differentiation in Drosophila. Development 125, 1361–1369 - PubMed
-
- Bate M., Rushton E., Currie D. A. (1991). Cells with persistent twist expression are the embryonic precursors of adult muscles in Drosophila. Development 113, 79–89 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
