

Epigenetic control of dendritic cell development and fate determination of common myeloid progenitor by Mysm1
- PMID: 25217698
- PMCID: PMC4208280
- DOI: 10.1182/blood-2013-10-534313
Epigenetic control of dendritic cell development and fate determination of common myeloid progenitor by Mysm1
Abstract
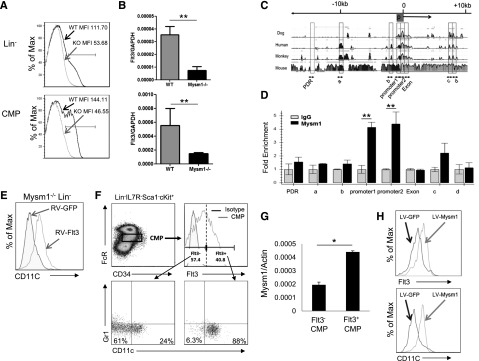
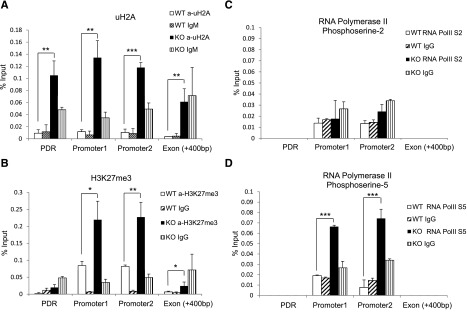
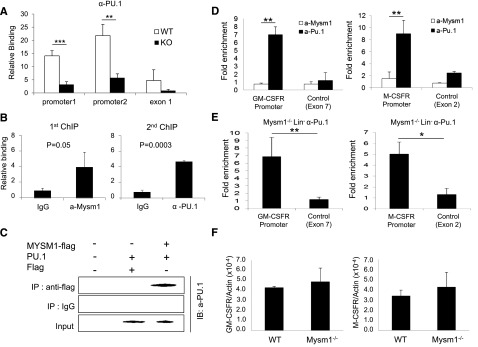
The mechanisms controlling the development of dendritic cells (DCs) remain incompletely understood. Using an Mysm1 knockout (Mysm1(-/-)) mouse model, we identified the histone H2A deubiquitinase Mysm1, as a critical regulator in DC differentiation. Mysm1(-/-) mice showed a global reduction of DCs in lymphoid organs, whereas development of granulocytes and macrophages were not severely affected. Hematopoietic progenitors and DC precursors were significantly decreased in Mysm1(-/-) mice and defective in Fms-like tyrosine kinase-3(Flt3) ligand-induced, but not in granulocyte macrophage-colony-stimulating factor (GM-CSF)-induced DC differentiation in vitro. Molecular studies demonstrated that the developmental defect of DCs from common myeloid progenitor (CMP) in Mysm1(-/-) mice is associated with decreased Flt3 expression and that Mysm1 derepresses transcription of the Flt3 gene by directing histone modifications at the Flt3 promoter region. Two molecular mechanisms were found to be responsible for the selective role of Mysm1 in lineage determination of DCs from CMPs: the selective expression of Mysm1 in a subset of CMPs and the different requirement of Mysm1 for PU.1 recruitment to the Flt3 locus vs GM-CSF-α and macrophage-colony-stimulating factor receptor loci. In conclusion, this study reveals an essential role of Mysm1 in epigenetic regulation of Flt3 transcription and DC development, and it provides a novel mechanism for lineage determination from CMP.
© 2014 by The American Society of Hematology.
Figures
Similar articles
-
The concerted action of GM-CSF and Flt3-ligand on in vivo dendritic cell homeostasis.Blood. 2009 Jul 23;114(4):835-43. doi: 10.1182/blood-2009-02-206318. Epub 2009 May 22. Blood. 2009. PMID: 19465690
-
Chromatin-binding deubiquitinase MYSM1 acts in haematopoietic progenitors to control dendritic cell development and to program dendritic cell responses to microbial stimulation.Immunology. 2024 May;172(1):109-126. doi: 10.1111/imm.13758. Epub 2024 Feb 5. Immunology. 2024. PMID: 38316548
-
PU.1 is essential for CD11c expression in CD8(+)/CD8(-) lymphoid and monocyte-derived dendritic cells during GM-CSF or FLT3L-induced differentiation.PLoS One. 2012;7(12):e52141. doi: 10.1371/journal.pone.0052141. Epub 2012 Dec 20. PLoS One. 2012. PMID: 23284905 Free PMC article.
-
Stem cell fate specification: role of master regulatory switch transcription factor PU.1 in differential hematopoiesis.Stem Cells Dev. 2005 Apr;14(2):140-52. doi: 10.1089/scd.2005.14.140. Stem Cells Dev. 2005. PMID: 15910240 Review.
-
Expression of FLT3 receptor and response to FLT3 ligand by leukemic cells.Leukemia. 1996 Apr;10(4):588-99. Leukemia. 1996. PMID: 8618433 Review.
Cited by
-
The Role of Deubiquitinating Enzymes in Hematopoiesis and Hematological Malignancies.Cancers (Basel). 2020 Apr 28;12(5):1103. doi: 10.3390/cancers12051103. Cancers (Basel). 2020. PMID: 32354135 Free PMC article. Review.
-
MYSM-1 suppresses migration and invasion in renal carcinoma through inhibiting epithelial-mesenchymal transition.Tumour Biol. 2016 Dec;37:15583–15591. doi: 10.1007/s13277-015-4138-z. Epub 2015 Sep 27. Tumour Biol. 2016. PMID: 26409454
-
Yangonin treats inflammatory osteoporosis by inhibiting the secretion of inflammatory factors and RANKL expression.Inflammopharmacology. 2022 Aug;30(4):1445-1458. doi: 10.1007/s10787-022-00985-1. Epub 2022 Apr 22. Inflammopharmacology. 2022. PMID: 35451724
-
Dendritic Cells and Cancer: From Biology to Therapeutic Intervention.Cancers (Basel). 2019 Apr 11;11(4):521. doi: 10.3390/cancers11040521. Cancers (Basel). 2019. PMID: 30979057 Free PMC article. Review.
-
Deubiquitinase MYSM1: An Important Tissue Development and Function Regulator.Int J Mol Sci. 2024 Dec 4;25(23):13051. doi: 10.3390/ijms252313051. Int J Mol Sci. 2024. PMID: 39684760 Free PMC article. Review.
References
-
- Schmid MA, Kingston D, Boddupalli S, Manz MG. Instructive cytokine signals in dendritic cell lineage commitment. Immunol Rev. 2010;234(1):32–44. - PubMed
-
- Ebner S, Ratzinger G, Krösbacher B, et al. Production of IL-12 by human monocyte-derived dendritic cells is optimal when the stimulus is given at the onset of maturation, and is further enhanced by IL-4. J Immunol. 2001;166(1):633–641. - PubMed
-
- Xu Y, Zhan Y, Lew AM, Naik SH, Kershaw MH. Differential development of murine dendritic cells by GM-CSF versus Flt3 ligand has implications for inflammation and trafficking. J Immunol. 2007;179(11):7577–7584. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
