

Volatile evolution of long noncoding RNA repertoires: mechanisms and biological implications
- PMID: 25218058
- PMCID: PMC4464757
- DOI: 10.1016/j.tig.2014.08.004
Volatile evolution of long noncoding RNA repertoires: mechanisms and biological implications
Abstract
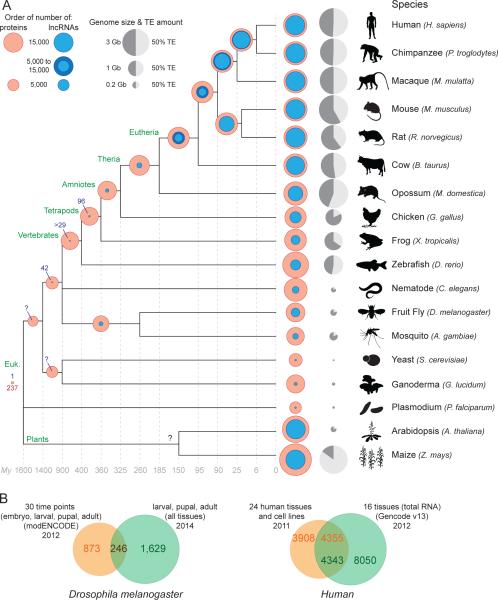
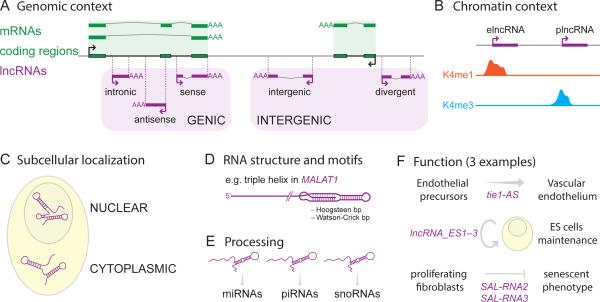
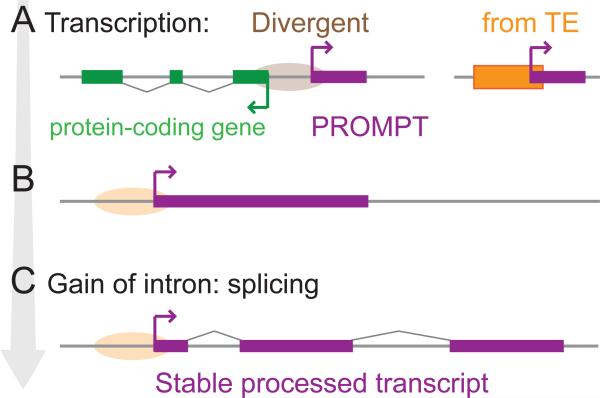
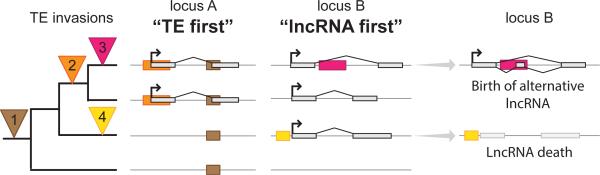
Thousands of genes encoding long noncoding RNAs (lncRNAs) have been identified in all vertebrate genomes thus far examined. The list of lncRNAs partaking in arguably important biochemical, cellular, and developmental activities is steadily growing. However, it is increasingly clear that lncRNA repertoires are subject to weak functional constraint and rapid turnover during vertebrate evolution. We discuss here some of the factors that may explain this apparent paradox, including relaxed constraint on sequence to maintain lncRNA structure/function, extensive redundancy in the regulatory circuits in which lncRNAs act, as well as adaptive and non-adaptive forces such as genetic drift. We explore the molecular mechanisms promoting the birth and rapid evolution of lncRNA genes, with an emphasis on the influence of bidirectional transcription and transposable elements, two pervasive features of vertebrate genomes. Together these properties reveal a remarkably dynamic and malleable noncoding transcriptome which may represent an important source of robustness and evolvability.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Necsulea A, et al. The evolution of lncRNA repertoires and expression patterns in tetrapods. Nature. 2014;505(7485):635–640. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
