

Biological messiness vs. biological genius: Mechanistic aspects and roles of protein promiscuity
- PMID: 25218442
- PMCID: PMC4920067
- DOI: 10.1016/j.jsbmb.2014.09.010
Biological messiness vs. biological genius: Mechanistic aspects and roles of protein promiscuity
Abstract
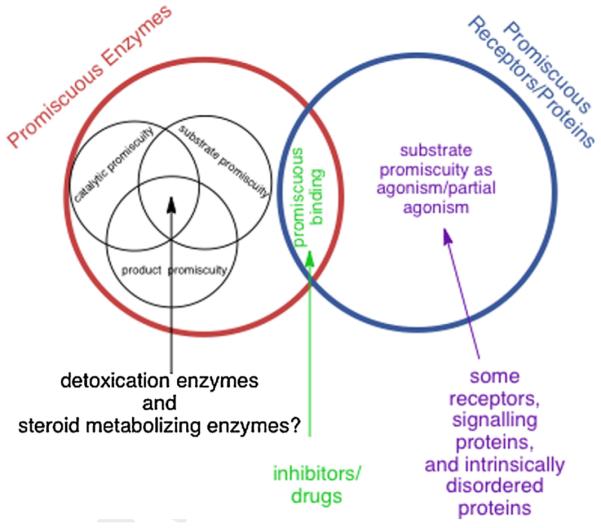
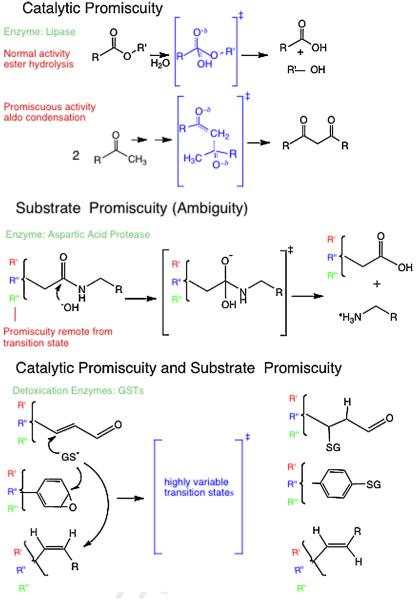
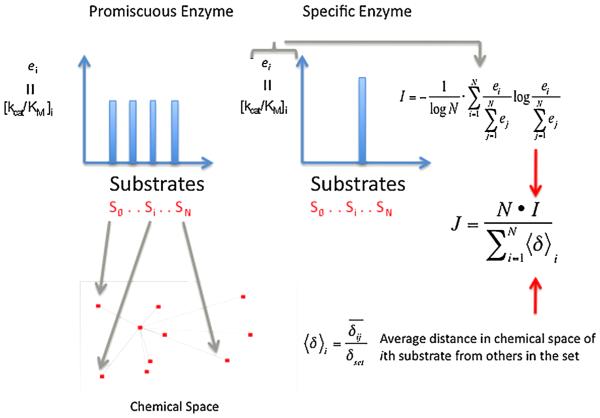
In contrast to the traditional biological paradigms focused on 'specificity', recent research and theoretical efforts have focused on functional 'promiscuity' exhibited by proteins and enzymes in many biological settings, including enzymatic detoxication, steroid biochemistry, signal transduction and immune responses. In addition, divergent evolutionary processes are apparently facilitated by random mutations that yield promiscuous enzyme intermediates. The intermediates, in turn, provide opportunities for further evolution to optimize new functions from existing protein scaffolds. In some cases, promiscuity may simply represent the inherent plasticity of proteins resulting from their polymeric nature with distributed conformational ensembles. Enzymes or proteins that bind or metabolize noncognate substrates create 'messiness' or noise in the systems they contribute to. With our increasing awareness of the frequency of these promiscuous behaviors it becomes interesting and important to understand the molecular bases for promiscuous behavior and to distinguish between evolutionarily selected promiscuity and evolutionarily tolerated messiness. This review provides an overview of current understanding of these aspects of protein biochemistry and enzymology.
Keywords: Detoxication enzymes; Enzyme evolution; Enzyme promiscuity; Evolvability; Intrinsically Disordered proteins.
Copyright © 2014. Published by Elsevier Ltd.
Figures
References
-
- Fersht A, Winter G. Protein engineering. Trends Biochem. Sci. 1992;17(8):292–295. - PubMed
-
- Russell AJ, Fersht AR. Rational modification of enzyme catalysis by engineering surface charge. Nature. 1987;328(6130):496–500. - PubMed
-
- Fersht AR, Shi JP, Knill-Jones J, Lowe DM, Wilkinson AJ, Blow DM, Brick P, Carter P, Waye MM, Winter G. Hydrogen bonding and biological specificity analysed by protein engineering. Nature. 1985;314(6008):235–238. - PubMed
-
- Sinha N, Smith-Gill SJ. Electrostatics in protein binding and function. Curr. Protein Pept. Sci. 2002;3(6):601–614. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
