

Distinct contributions of Aire and antigen-presenting-cell subsets to the generation of self-tolerance in the thymus
- PMID: 25220213
- PMCID: PMC4175925
- DOI: 10.1016/j.immuni.2014.08.007
Distinct contributions of Aire and antigen-presenting-cell subsets to the generation of self-tolerance in the thymus
Abstract
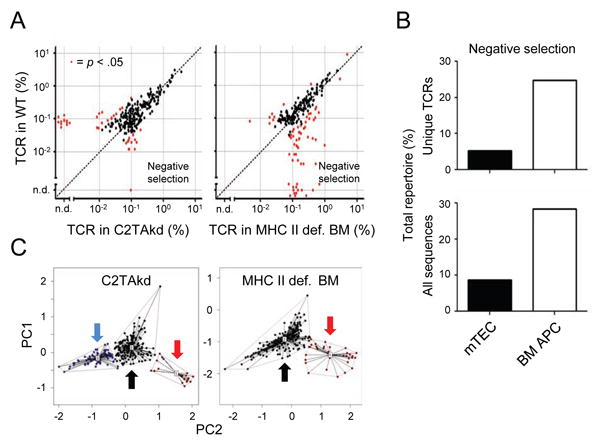
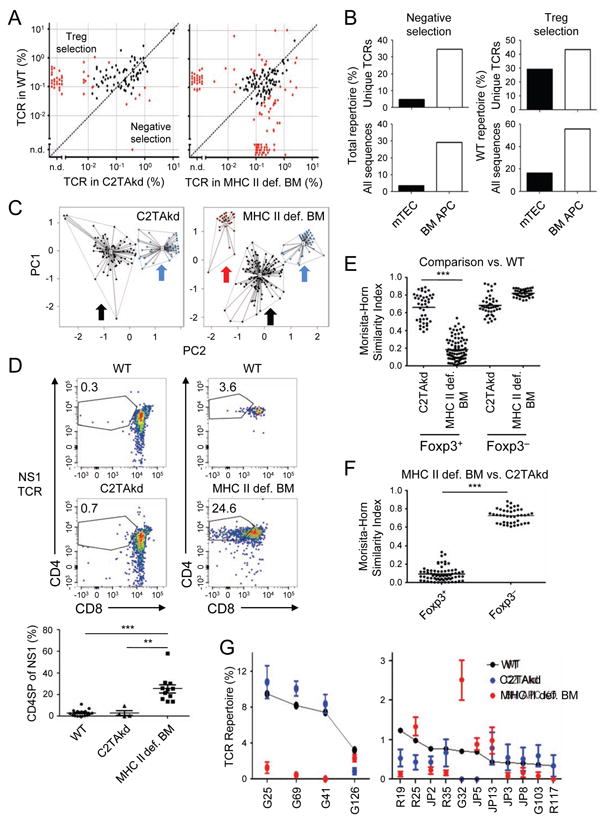
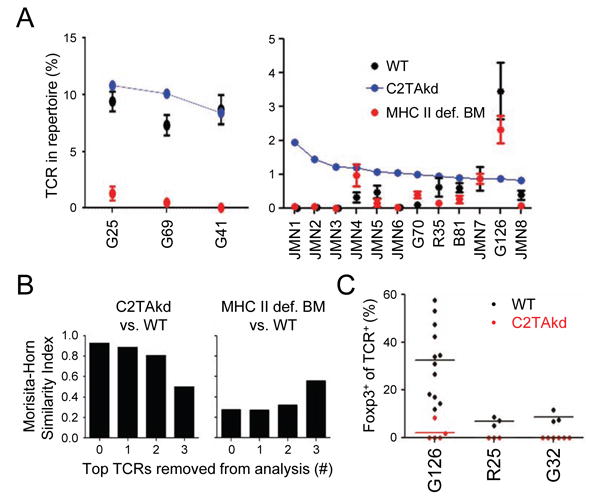
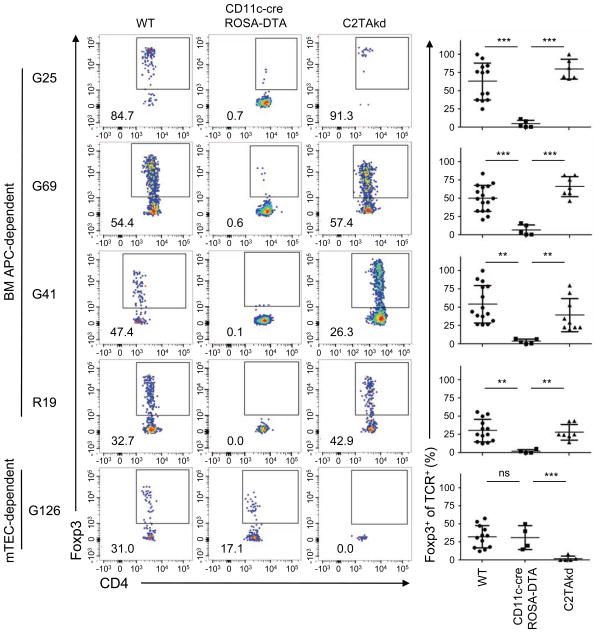
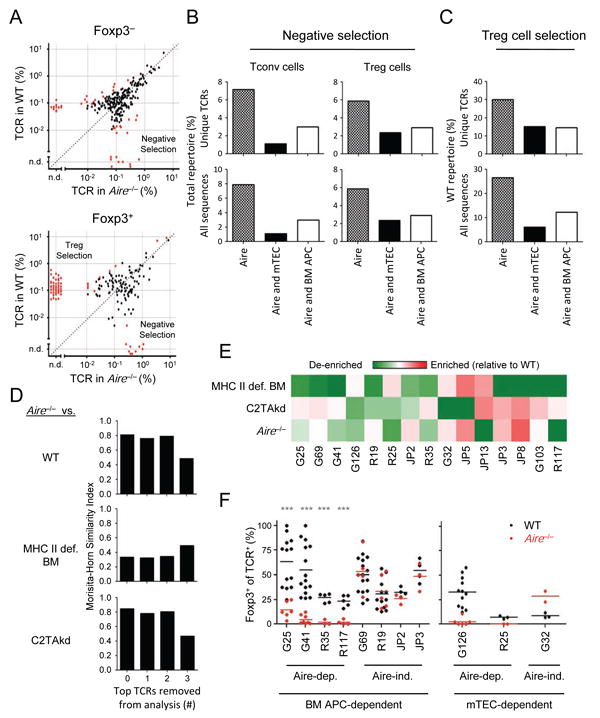
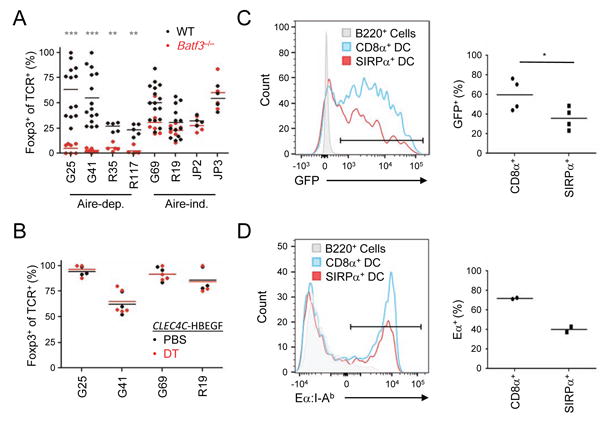
The contribution of thymic antigen-presenting-cell (APC) subsets in selecting a self-tolerant T cell population remains unclear. We show that bone marrow (BM) APCs and medullary thymic epithelial cells (mTECs) played nonoverlapping roles in shaping the T cell receptor (TCR) repertoire by deletion and regulatory T (Treg) cell selection of distinct TCRs. Aire, which induces tissue-specific antigen expression in mTECs, affected the TCR repertoire in a manner distinct from mTEC presentation. Approximately half of Aire-dependent deletion or Treg cell selection utilized a pathway dependent on antigen presentation by BM APCs. Batf3-dependent CD8α⁺ dendritic cells (DCs) were the crucial BM APCs for Treg cell selection via this pathway, showing enhanced ability to present antigens from stromal cells. These results demonstrate the division of function between thymic APCs in shaping the self-tolerant TCR repertoire and reveal an unappreciated cooperation between mTECs and CD8α⁺ DCs for presentation of Aire-induced self-antigens to developing thymocytes.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Love is in the Aire: mTECs share their assets.Immunity. 2014 Sep 18;41(3):343-345. doi: 10.1016/j.immuni.2014.08.013. Immunity. 2014. PMID: 25238088
-
Neonatal immunity: babies' T cells can fight.Nat Rev Immunol. 2014 Nov;14(11):714-5. doi: 10.1038/nri3758. Epub 2014 Oct 10. Nat Rev Immunol. 2014. PMID: 25301253 No abstract available.
References
-
- Anderson MS, Venanzi ES, Chen Z, Berzins SP, Benoist C, Mathis D. The Cellular Mechanism of Aire Control of T Cell Tolerance. Immunity. 2005;23:227–239. - PubMed
-
- Aschenbrenner K, D'Cruz LM, Vollmann EH, Hinterberger M, Emmerich J, Swee LK, Rolink A, Klein L. Selection of Foxp3+ regulatory T cells specific for self antigen expressed and presented by Aire+ medullary thymic epithelial cells. Nat Immunol. 2007;8:351–358. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
