

Inward rectifiers and their regulation by endogenous polyamines
- PMID: 25221519
- PMCID: PMC4145359
- DOI: 10.3389/fphys.2014.00325
Inward rectifiers and their regulation by endogenous polyamines
Abstract
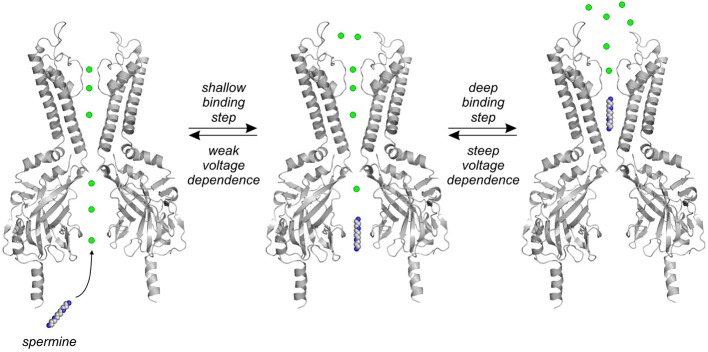
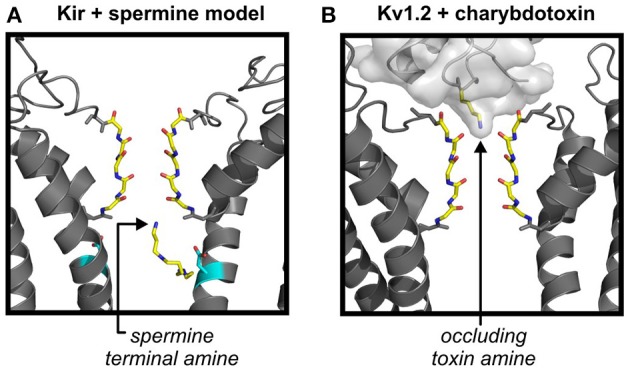
Inwardly-rectifying potassium (Kir) channels contribute to maintenance of the resting membrane potential and regulation of electrical excitation in many cell types. Strongly rectifying Kir channels exhibit a very steep voltage dependence resulting in silencing of their activity at depolarized membrane voltages. The mechanism underlying this steep voltage dependence is blockade by endogenous polyamines. These small multifunctional, polyvalent metabolites enter the long Kir channel pore from the intracellular side, displacing multiple occupant ions as they migrate to a stable binding site in the transmembrane region of the channel. Numerous structure-function studies have revealed structural elements of Kir channels that determine their susceptibility to polyamine block, and enable the steep voltage dependence of this process. In addition, various channelopathies have been described that result from alteration of the polyamine sensitivity or activity of strongly rectifying channels. The primary focus of this article is to summarize current knowledge of the molecular mechanisms of polyamine block, and provide some perspective on lingering uncertainties related to this physiologically important mechanism of ion channel blockade. We also briefly review some of the important and well understood physiological roles of polyamine sensitive, strongly rectifying Kir channels, primarily of the Kir2 family.
Keywords: channelopathy; inward rectifier; ion channel block; polyamines; potassium channels; voltage-dependent gating.
Figures




References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources