

Endothelial dysfunction is a potential contributor to multiple organ failure and mortality in aged mice subjected to septic shock: preclinical studies in a murine model of cecal ligation and puncture
- PMID: 25223540
- PMCID: PMC4177582
- DOI: 10.1186/s13054-014-0511-3
Endothelial dysfunction is a potential contributor to multiple organ failure and mortality in aged mice subjected to septic shock: preclinical studies in a murine model of cecal ligation and puncture
Abstract
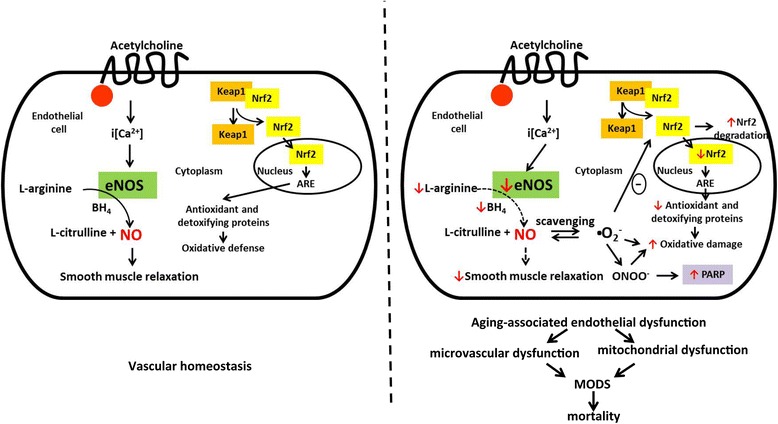
Introduction: The goal of the current study was to investigate the effect of aging on the development of endothelial dysfunction in a murine model of sepsis, and to compare it with the effect of genetic deficiency of the endothelial isoform of nitric oxide synthase (eNOS).
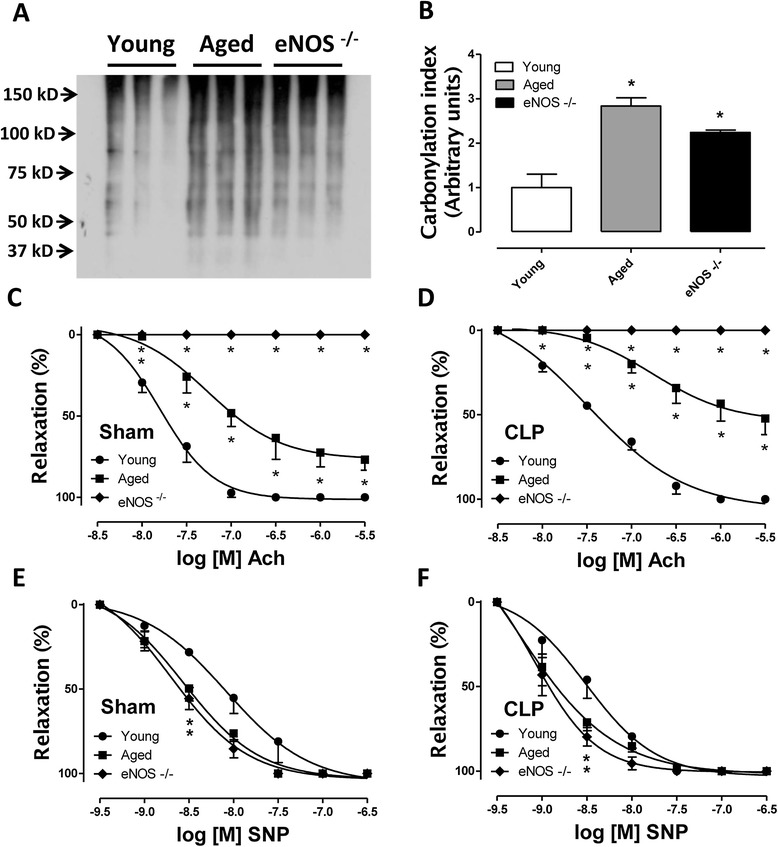
Methods: Cecal ligation and puncture (CLP) was used to induce sepsis in mice. Survival rates were monitored and plasma indices of organ function were measured. Ex vivo studies included the measurement of vascular function in thoracic aortic rings, assessment of oxidative stress/cellular injury in various organs and the measurement of mitochondrial function in isolated liver mitochondria.
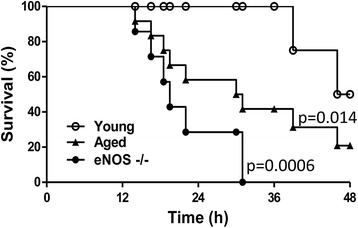
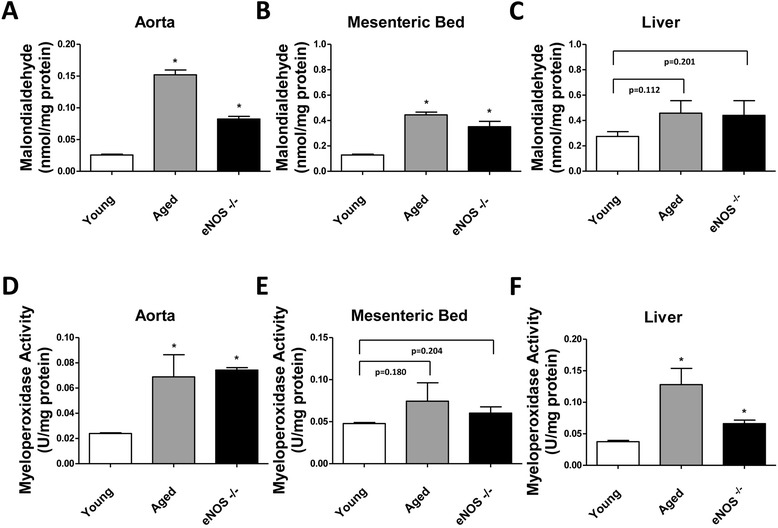
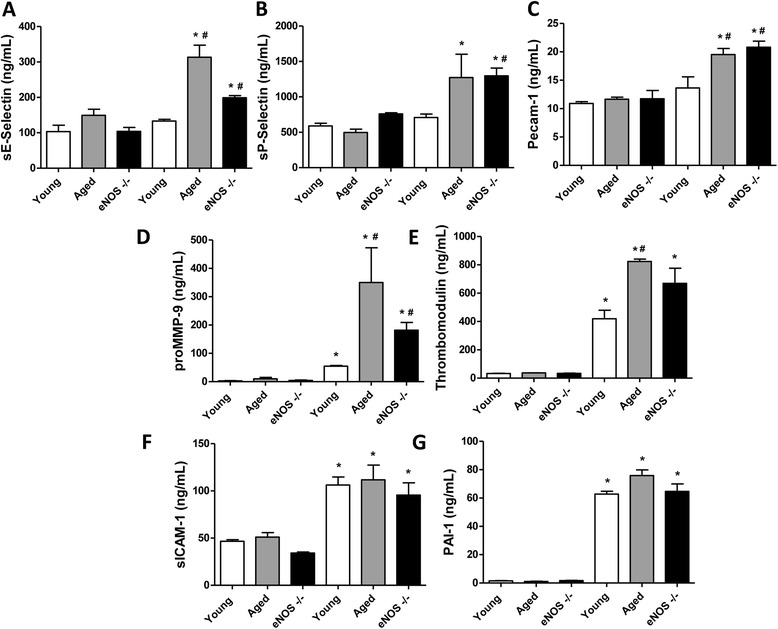
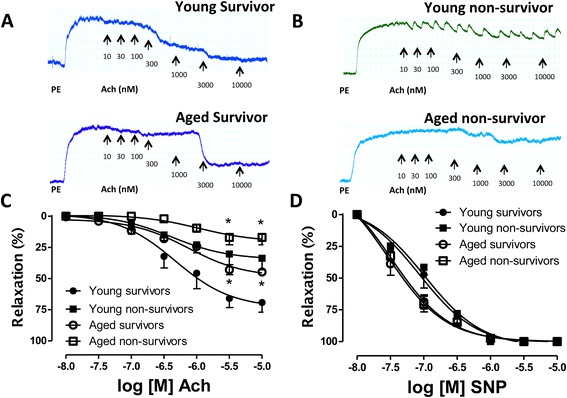
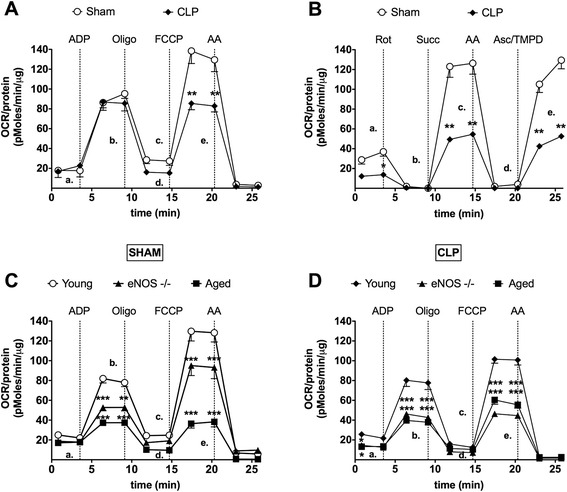
Results: eNOS deficiency and aging both exacerbated the mortality of sepsis. Both eNOS-deficient and aged mice exhibited a higher degree of sepsis-associated multiple organ dysfunction syndrome (MODS), infiltration of tissues with mononuclear cells and oxidative stress. A high degree of sepsis-induced vascular oxidative damage and endothelial dysfunction (evidenced by functional assays and multiple plasma markers of endothelial dysfunction) was detected in aortae isolated from both eNOS(-/-) and aged mice. There was a significant worsening of sepsis-induced mitochondrial dysfunction, both in eNOS-deficient mice and in aged mice. Comparison of the surviving and non-surviving groups of animals indicated that the severity of endothelial dysfunction may be a predictor of mortality of mice subjected to CLP-induced sepsis.
Conclusions: Based on the studies in eNOS mice, we conclude that the lack of endothelial nitric oxide production, on its own, may be sufficient to markedly exacerbate the severity of septic shock. Aging markedly worsens the degree of endothelial dysfunction in sepsis, yielding a significant worsening of the overall outcome. Thus, endothelial dysfunction may constitute an early predictor and independent contributor to sepsis-associated MODS and mortality in aged mice.
Figures
References
-
- Martin GS, Mannino DM, Moss M. The effect of age on the development and outcome of adult sepsis. Crit Care Med. 2006;34:15–21. doi: 10.1097/01.CCM.0000194535.82812.BA. - DOI - PubMed
-
- Altura BM, Gebrewold A, Burton RW. Failure of microscopic metarterioles to elicit vasodilator responses to acetylcholine, bradykinin, histamine and substance P after ischemic shock, endotoxemia and trauma: possible role of endothelial cells. Microcirc Endothelium Lymphatics. 1985;2:121–127. - PubMed
-
- Szabo C, Farago M, Horvath I, Lohinai Z, Kovach AG. Hemorrhagic hypotension impairs endothelium-dependent relaxations in the renal artery of the cat. Circ Shock. 1992;36:238–241. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
