

Comparison of TALE designer transcription factors and the CRISPR/dCas9 in regulation of gene expression by targeting enhancers
- PMID: 25223790
- PMCID: PMC4227760
- DOI: 10.1093/nar/gku836
Comparison of TALE designer transcription factors and the CRISPR/dCas9 in regulation of gene expression by targeting enhancers
Abstract
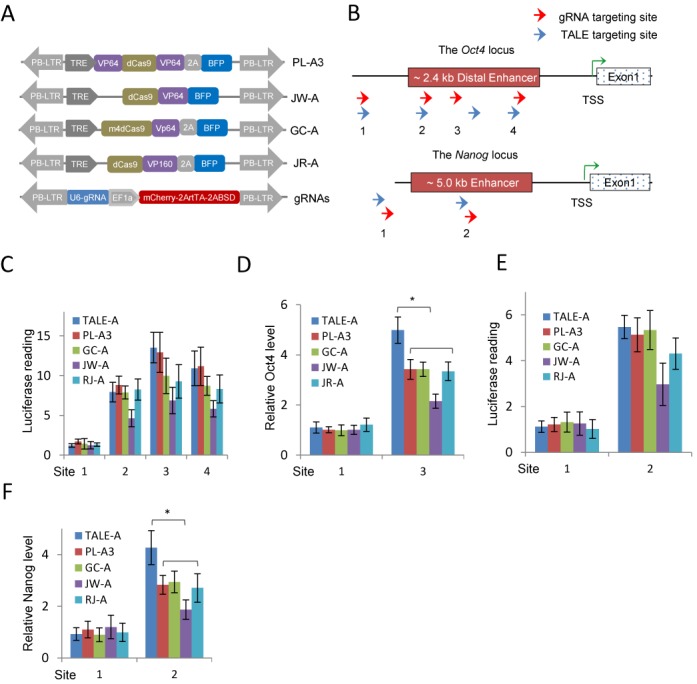
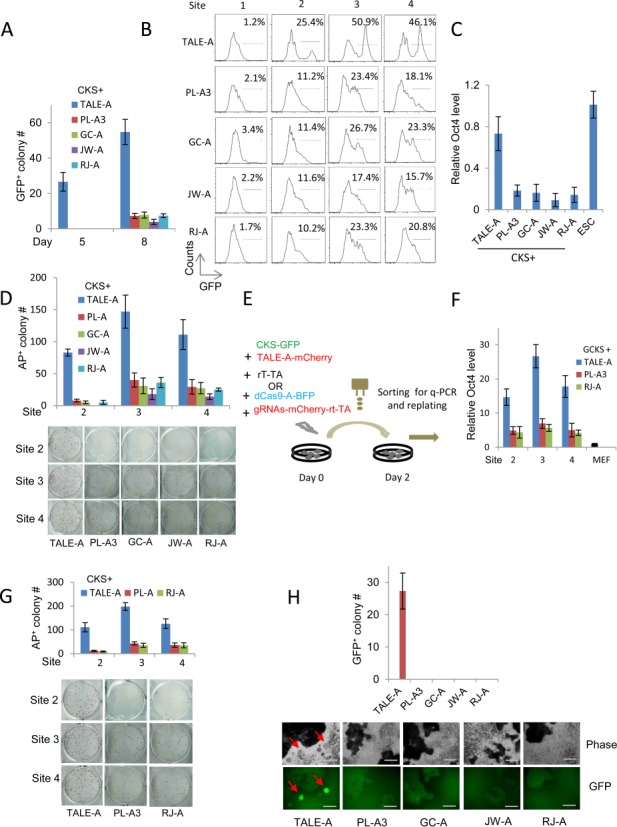
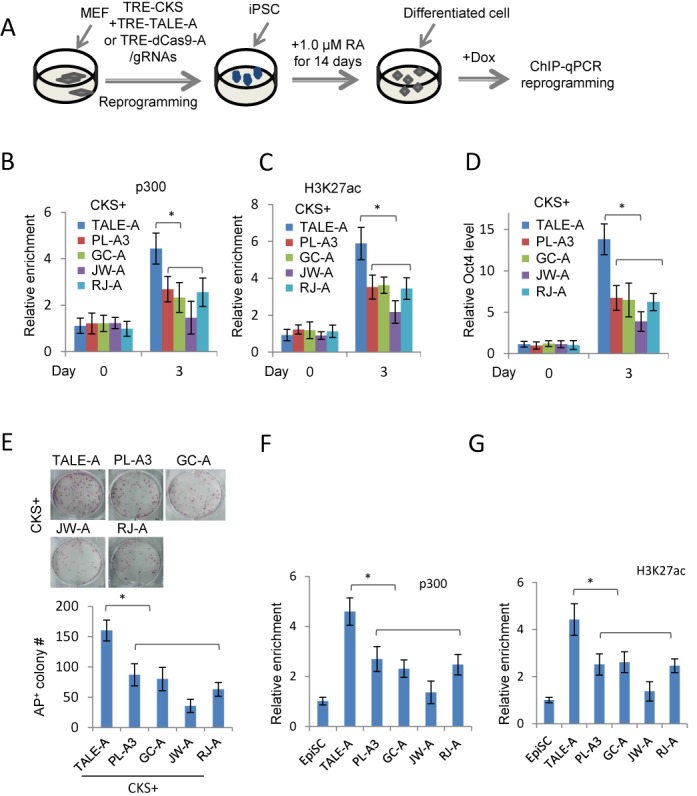
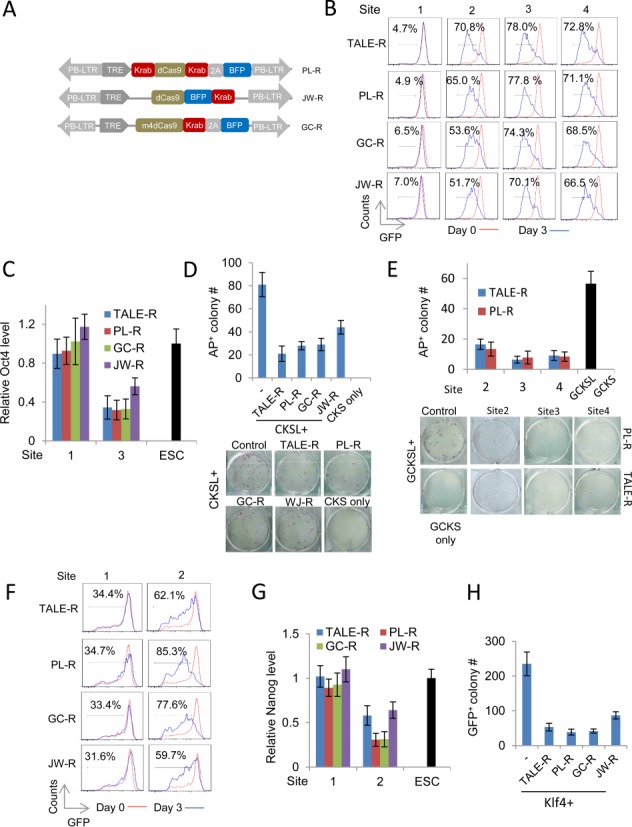
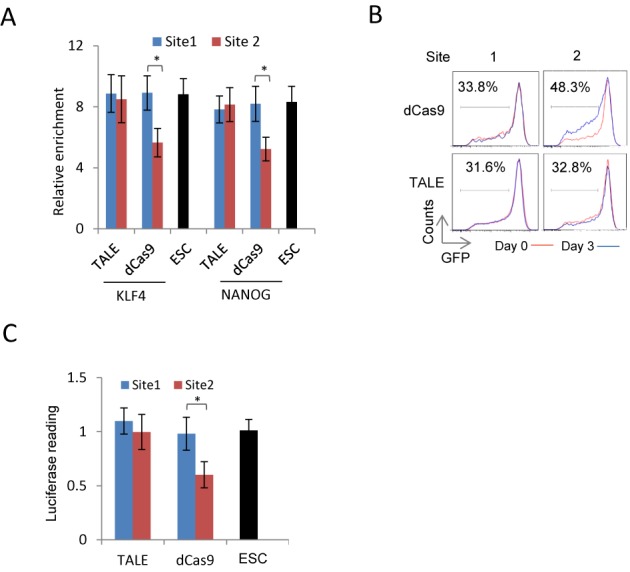
The transcription activator-like effectors (TALEs) and the RNA-guided clustered regularly interspaced short palindromic repeat (CRISPR) associated protein (Cas9) utlilize distinct molecular mechanisms in targeting site recognition. The two proteins can be modified to carry additional functional domains to regulate expression of genomic loci in mammalian cells. In this study, we have compared the two systems in activation and suppression of the Oct4 and Nanog loci by targeting their enhancers. Although both are able to efficiently activate the luciferase reporters, the CRISPR/dCas9 system is much less potent in activating the endogenous loci and in the application of reprogramming somatic cells to iPS cells. Nevertheless, repression by CRISPR/dCas9 is comparable to or even better than TALE repressors. We demonstrated that dCas9 protein binding results in significant physical interference to binding of native transcription factors at enhancer, less efficient active histone markers induction or recruitment of activating complexes in gene activation. This study thus highlighted the merits and drawbacks of transcription regulation by each system. A combined approach of TALEs and CRISPR/dCas9 should provide an optimized solution to regulate genomic loci and to study genetic elements such as enhancers in biological processes including somatic cell reprogramming and guided differentiation.
© The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Takahashi K., Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. - PubMed
-
- Brons I.G., Smithers L.E., Trotter M.W., Rugg-Gunn P., Sun B., Chuva de Sousa Lopes S.M., Howlett S.K., Clarkson A., Ahrlund-Richter L., Pedersen R.A., et al. Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature. 2007;448:191–195. - PubMed
-
- Li R., Liang J., Ni S., Zhou T., Qing X., Li H., He W., Chen J., Li F., Zhuang Q., et al. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell. 2010;7:51–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
