

Charge requirements of lipid II flippase activity in Escherichia coli
- PMID: 25225268
- PMCID: PMC4248868
- DOI: 10.1128/JB.02172-14
Charge requirements of lipid II flippase activity in Escherichia coli
Abstract
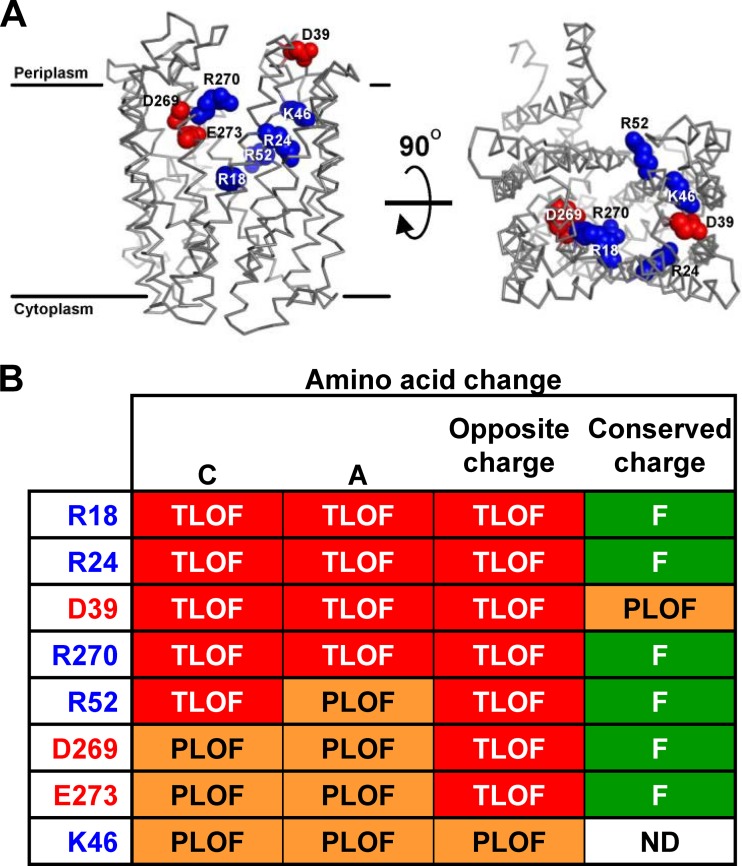
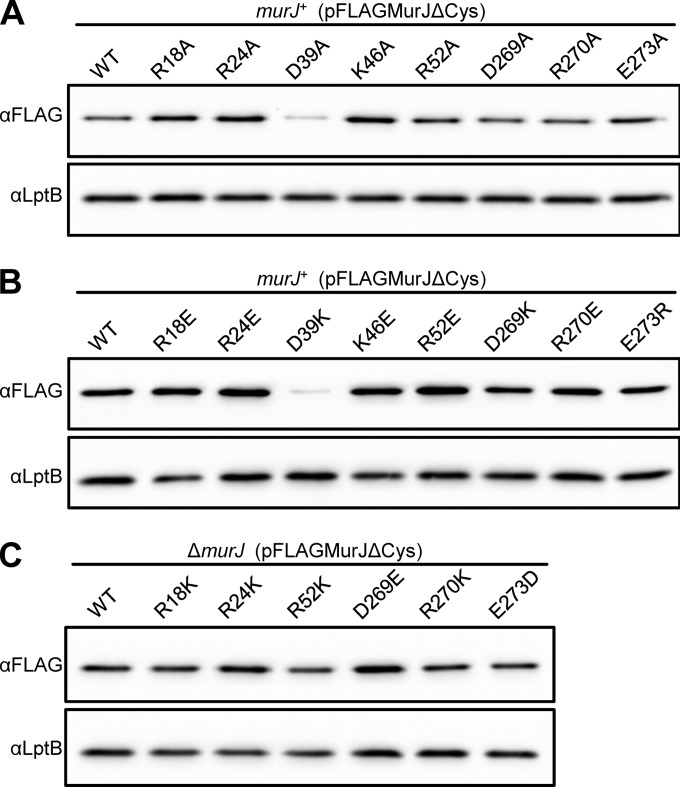
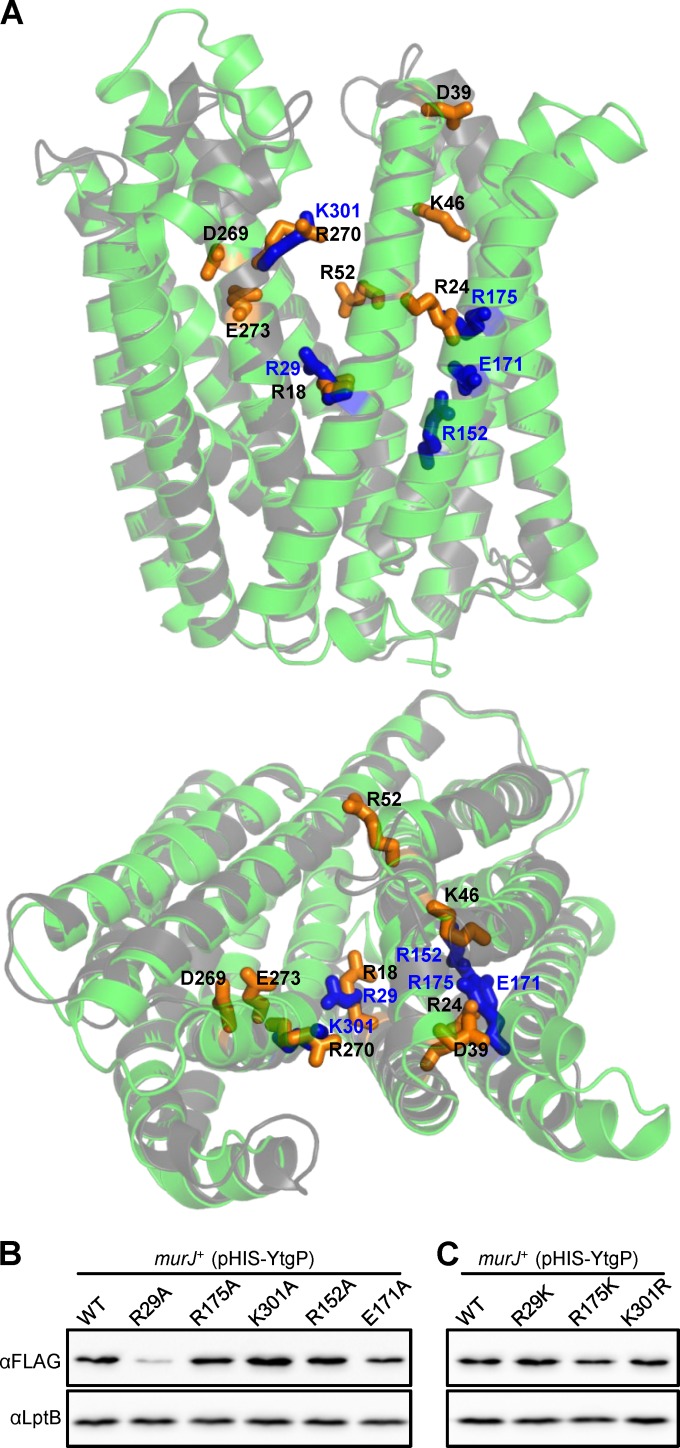
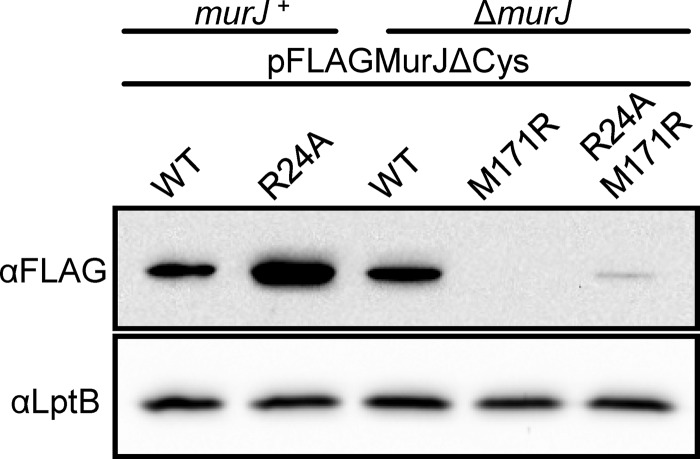
Peptidoglycan (PG) is an extracytoplasmic glycopeptide matrix essential for the integrity of the envelope of most bacteria. The PG building block is a disaccharide-pentapeptide that is synthesized as a lipid-linked precursor called lipid II. The translocation of the amphipathic lipid II across the cytoplasmic membrane is required for subsequent incorporation of the disaccharide-pentapeptide into PG. In Escherichia coli, the essential inner membrane protein MurJ is the lipid II flippase. Previous studies showed that 8 charged residues in the central cavity region of MurJ are crucial for function. Here, we completed the functional analysis of all 57 charged residues in MurJ and demonstrated that the respective positive or negative charge of the 8 aforementioned residues is required for proper MurJ function. Loss of the negative charge in one of these residues, D39, causes a severe defect in MurJ biogenesis; by engineering an intragenic suppressor mutation that restores MurJ biogenesis, we found that this charge is also essential for MurJ function. Because of the low level of homology between MurJ and putative orthologs from Gram-positive bacteria, we explored the conservation of these 8 charged residues in YtgP, a homolog from Streptococcus pyogenes. We found that only 3 positive charges are similarly positioned and essential in YtgP; YtgP possesses additional charged residues within its predicted cavity that are essential for function and conserved among Gram-positive bacteria. From these data, we hypothesize that some charged residues in the cavity region of MurJ homologs are required for interaction with lipid II and/or energy coupling during transport.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
