

Higher brain functions served by the lowly rodent primary visual cortex
- PMID: 25225298
- PMCID: PMC4175492
- DOI: 10.1101/lm.034355.114
Higher brain functions served by the lowly rodent primary visual cortex
Abstract
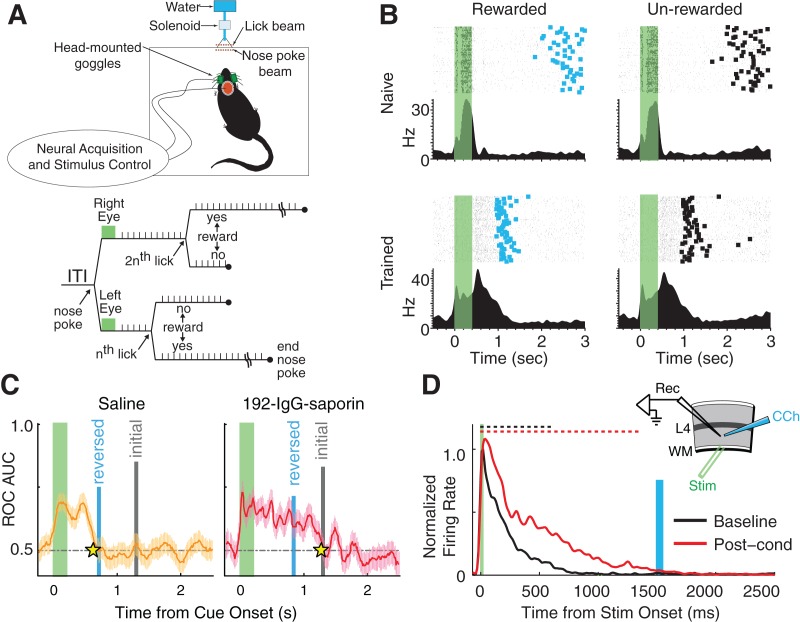
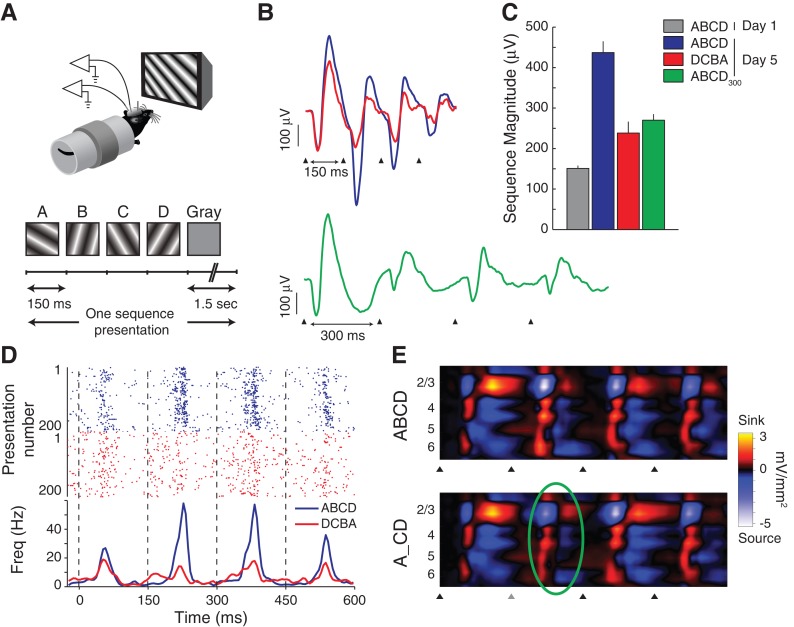
It has been more than 50 years since the first description of ocular dominance plasticity--the profound modification of primary visual cortex (V1) following temporary monocular deprivation. This discovery immediately attracted the intense interest of neurobiologists focused on the general question of how experience and deprivation modify the brain as a potential substrate for learning and memory. The pace of discovery has quickened considerably in recent years as mice have become the preferred species to study visual cortical plasticity, and new studies have overturned the dogma that primary sensory cortex is immutable after a developmental critical period. Recent work has shown that, in addition to ocular dominance plasticity, adult visual cortex exhibits several forms of response modification previously considered the exclusive province of higher cortical areas. These "higher brain functions" include neural reports of stimulus familiarity, reward-timing prediction, and spatiotemporal sequence learning. Primary visual cortex can no longer be viewed as a simple visual feature detector with static properties determined during early development. Rodent V1 is a rich and dynamic cortical area in which functions normally associated only with "higher" brain regions can be studied at the mechanistic level.
© 2014 Gavornik and Bear; Published by Cold Spring Harbor Laboratory Press.
Figures

References
-
- Bach-y-Rita P, Kercel SW 2003. Sensory substitution and the human-machine interface. Trends Cogn Sci 7: 541–546 - PubMed
-
- Balas B, Sinha P 2008. Observing object motion induces increased generalization and sensitivity. Perception 37: 1160–1174 - PubMed
-
- Balas BJ, Sinha P 2009. The role of sequence order in determining view canonicality for novel wire-frame objects. Atten Percept Psychophys 71: 712–723 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources