

Probing membrane protein structure using water polarization transfer solid-state NMR
- PMID: 25228502
- PMCID: PMC4398059
- DOI: 10.1016/j.jmr.2014.08.007
Probing membrane protein structure using water polarization transfer solid-state NMR
Abstract
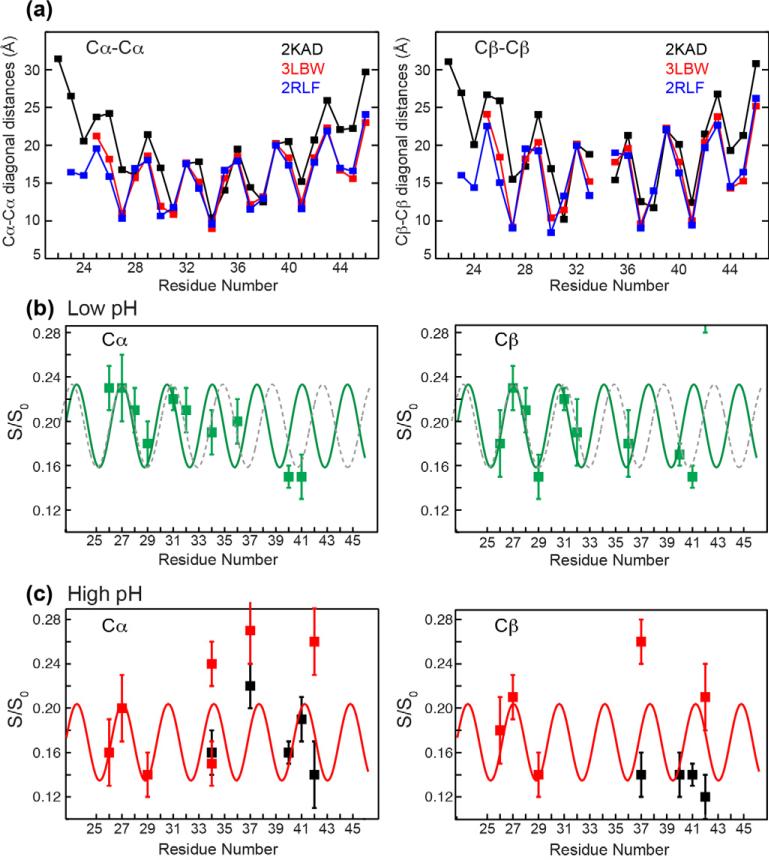
Water plays an essential role in the structure and function of proteins, lipid membranes and other biological macromolecules. Solid-state NMR heteronuclear-detected (1)H polarization transfer from water to biomolecules is a versatile approach for studying water-protein, water-membrane, and water-carbohydrate interactions in biology. We review radiofrequency pulse sequences for measuring water polarization transfer to biomolecules, the mechanisms of polarization transfer, and the application of this method to various biological systems. Three polarization transfer mechanisms, chemical exchange, spin diffusion and NOE, manifest themselves at different temperatures, magic-angle-spinning frequencies, and pulse irradiations. Chemical exchange is ubiquitous in all systems examined so far, and spin diffusion plays the key role in polarization transfer within the macromolecule. Tightly bound water molecules with long residence times are rare in proteins at ambient temperature. The water polarization-transfer technique has been used to study the hydration of microcrystalline proteins, lipid membranes, and plant cell wall polysaccharides, and to derive atomic-resolution details of the kinetics and mechanism of ion conduction in channels and pumps. Using this approach, we have measured the water polarization transfer to the transmembrane domain of the influenza M2 protein to obtain information on the structure of this tetrameric proton channel. At short mixing times, the polarization transfer rates are site-specific and depend on the pH, labile protons, sidechain conformation, as well as the radial position of the residues in this four-helix bundle. Despite the multiple dependences, the initial transfer rates reflect the periodic nature of the residue positions from the water-filled pore, thus this technique provides a way of gleaning secondary structure information, helix tilt angle, and the oligomeric structure of membrane proteins.
Keywords: Chemical exchange; Heteronuclear correlation; Influenza M2 protein; Ion channels; Spin diffusion.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures






References
-
- Otting G, Liepinsh E, Wüthrich K. Protein hydration in aqueous solution. Science. 1991;254:974–980. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
