

Dual modes of CLOCK:BMAL1 inhibition mediated by Cryptochrome and Period proteins in the mammalian circadian clock
- PMID: 25228643
- PMCID: PMC4173159
- DOI: 10.1101/gad.249417.114
Dual modes of CLOCK:BMAL1 inhibition mediated by Cryptochrome and Period proteins in the mammalian circadian clock
Abstract
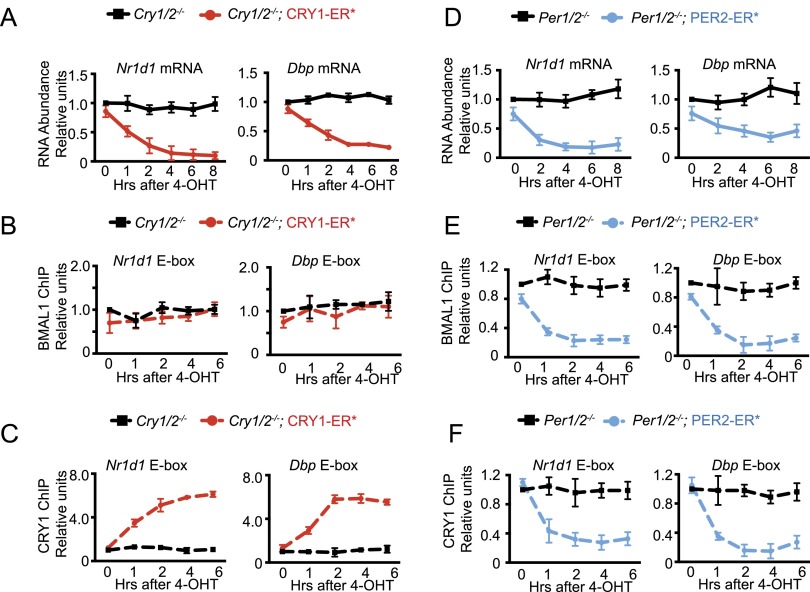
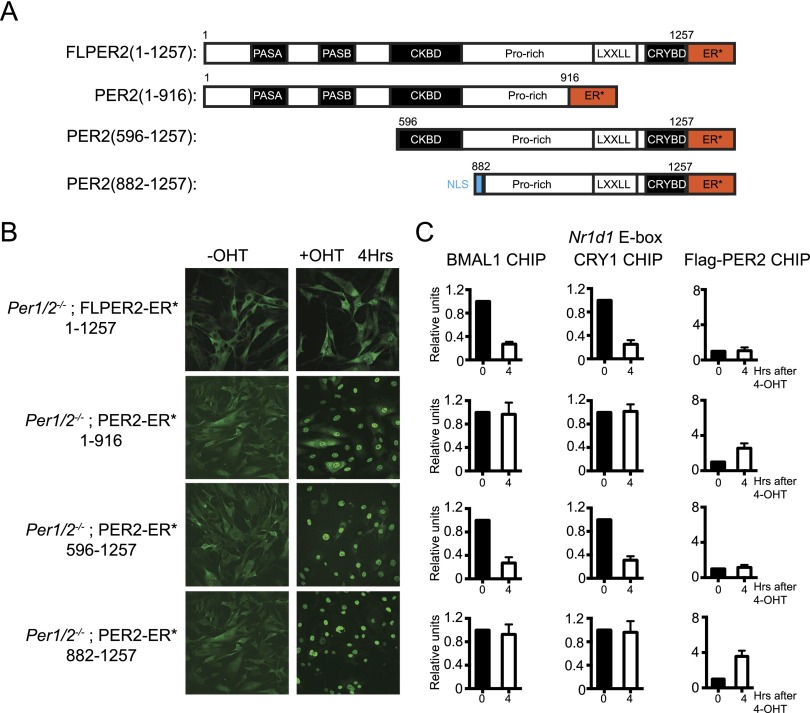
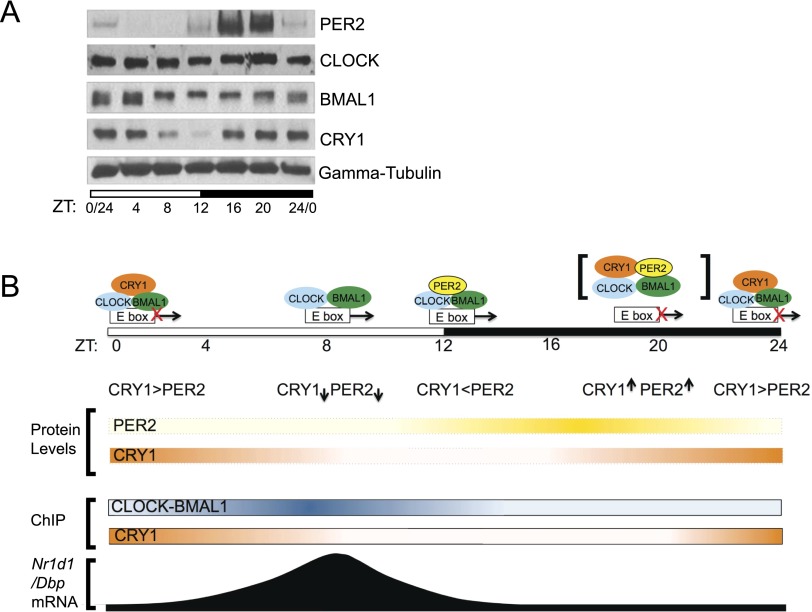
The mammalian circadian clock is based on a transcription-translation feedback loop (TTFL) in which CLOCK and BMAL1 proteins act as transcriptional activators of Cryptochrome and Period genes, which encode proteins that repress CLOCK-BMAL1 with a periodicity of ∼ 24 h. In this model, the mechanistic roles of CRY and PER are unclear. Here, we used a controlled targeting system to introduce CRY1 or PER2 into the nuclei of mouse cells with defined circadian genotypes to characterize the functions of CRY and PER. Our data show that CRY is the primary repressor in the TTFL: It binds to CLOCK-BMAL1 at the promoter and inhibits CLOCK-BMAL1-dependent transcription without dissociating the complex ("blocking"-type repression). PER alone has no effect on CLOCK-BMAL1-activated transcription. However, in the presence of CRY, nuclear entry of PER inhibits transcription by displacing CLOCK-BMAL1 from the promoter ("displacement"-type repression). In light of these findings, we propose a new model for the mammalian circadian clock in which the negative arm of the TTFL proceeds by two different mechanisms during the circadian cycle.
Keywords: Cryptochrome; DNA binding; Period; circadian; transcription.
© 2014 Ye et al.; Published by Cold Spring Harbor Laboratory Press.
Figures


References
-
- Albrecht U, Bordon A, Schmutz I, Ripperger J. 2007. The multiple facets of Per2. Cold Spring Harb Symp Quant Biol 72: 95–104 - PubMed
-
- Bae K, Jin X, Maywood ES, Hastings MH, Reppert SM, Weaver DR. 2001. Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock. Neuron 30: 525–536 - PubMed
-
- Brown SA, Ripperger J, Kadener S, Fleury-Olela F, Vilbois F, Rosbash M, Schibler U. 2005. PERIOD1-associated proteins modulate the negative limb of the mammalian circadian oscillator. Science 308: 693–696 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials