

Calcium entry into stereocilia drives adaptation of the mechanoelectrical transducer current of mammalian cochlear hair cells
- PMID: 25228765
- PMCID: PMC4205606
- DOI: 10.1073/pnas.1409920111
Calcium entry into stereocilia drives adaptation of the mechanoelectrical transducer current of mammalian cochlear hair cells
Abstract
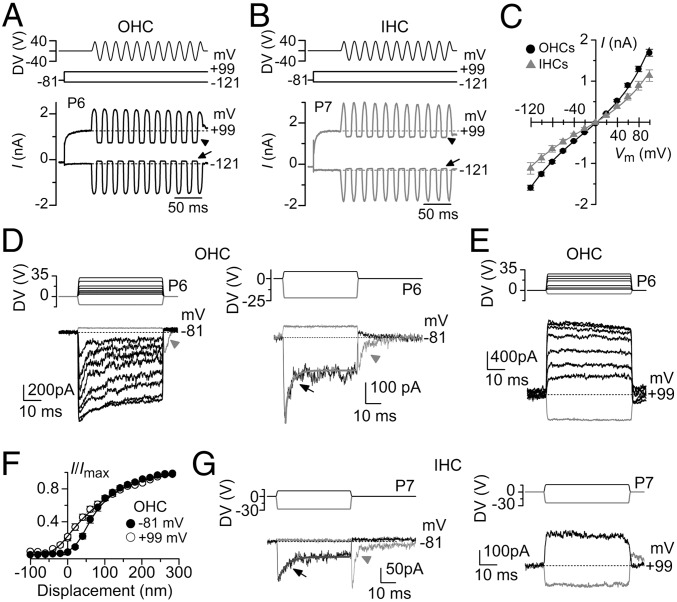
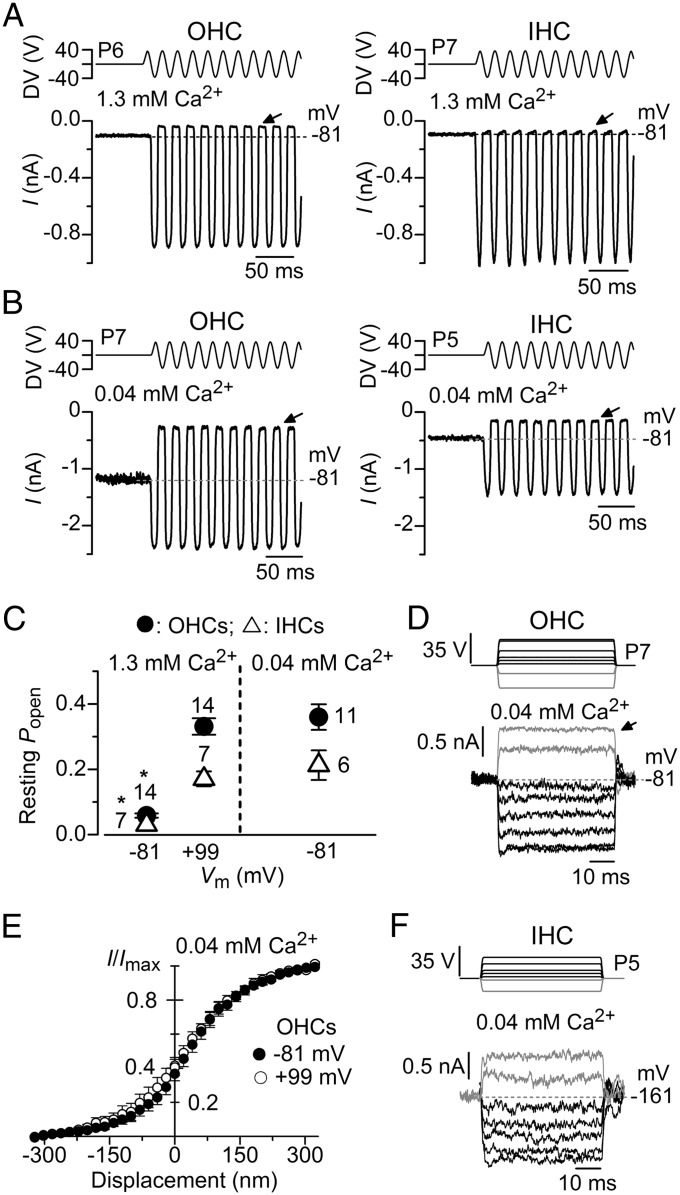
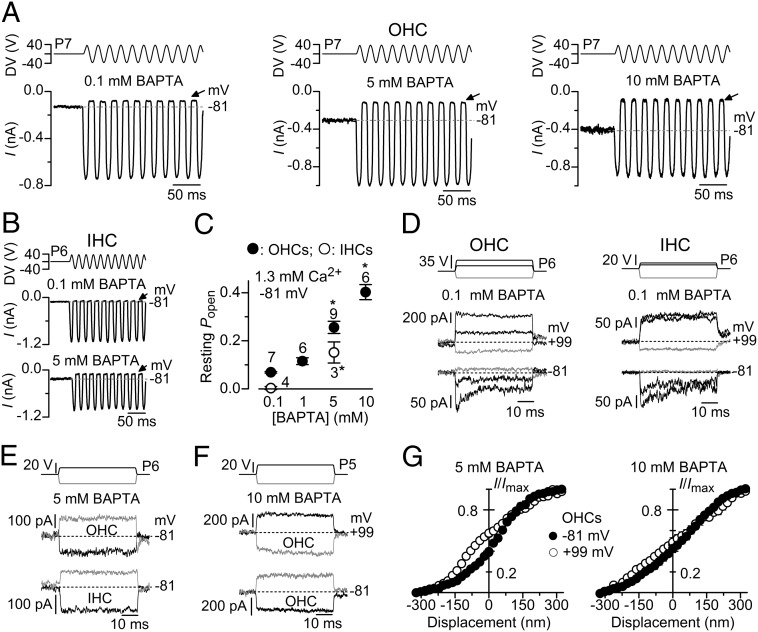
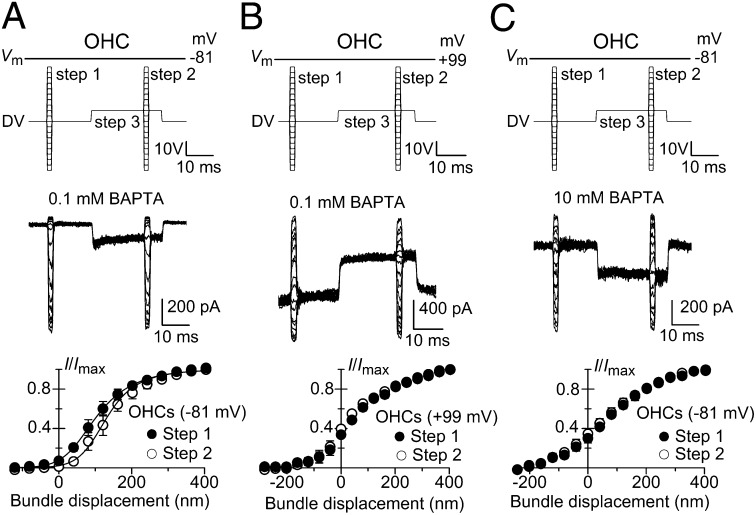
Mechanotransduction in the auditory and vestibular systems depends on mechanosensitive ion channels in the stereociliary bundles that project from the apical surface of the sensory hair cells. In lower vertebrates, when the mechanoelectrical transducer (MET) channels are opened by movement of the bundle in the excitatory direction, Ca(2+) entry through the open MET channels causes adaptation, rapidly reducing their open probability and resetting their operating range. It remains uncertain whether such Ca(2+)-dependent adaptation is also present in mammalian hair cells. Hair bundles of both outer and inner hair cells from mice were deflected by using sinewave or step mechanical stimuli applied using a piezo-driven fluid jet. We found that when cochlear hair cells were depolarized near the Ca(2+) reversal potential or their hair bundles were exposed to the in vivo endolymphatic Ca(2+) concentration (40 µM), all manifestations of adaptation, including the rapid decline of the MET current and the reduction of the available resting MET current, were abolished. MET channel adaptation was also reduced or removed when the intracellular Ca(2+) buffer 1,2-Bis(2-aminophenoxy)ethane-N,N,N',N'-tetraacetic acid (BAPTA) was increased from a concentration of 0.1 to 10 mM. The findings show that MET current adaptation in mouse auditory hair cells is modulated similarly by extracellular Ca(2+), intracellular Ca(2+) buffering, and membrane potential, by their common effect on intracellular free Ca(2+).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Fettiplace R, Hackney CM. The sensory and motor roles of auditory hair cells. Nat Rev Neurosci. 2006;7(1):19–29. - PubMed
-
- Pickles JO, et al. The organization of tip links and stereocilia on hair cells of bird and lizard basilar papillae. Hear Res. 1989;41(1):31–41. - PubMed
-
- Tilney LG, Tilney MS, DeRosier DJ. Actin filaments, stereocilia, and hair cells: How cells count and measure. Annu Rev Cell Biol. 1992;8:257–274. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
