

A cholesterol recognition motif in human phospholipid scramblase 1
- PMID: 25229146
- PMCID: PMC4167295
- DOI: 10.1016/j.bpj.2014.07.039
A cholesterol recognition motif in human phospholipid scramblase 1
Abstract
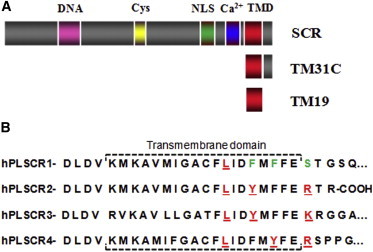
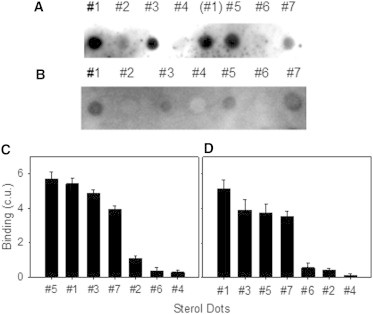
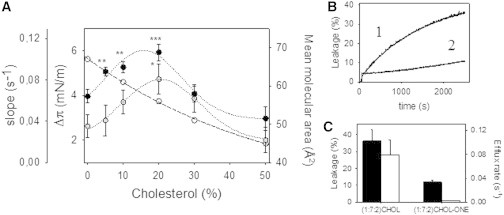
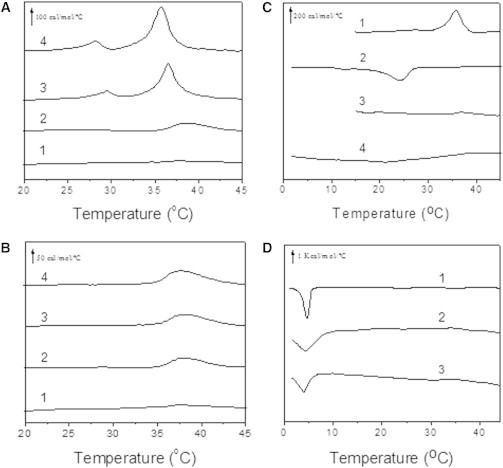
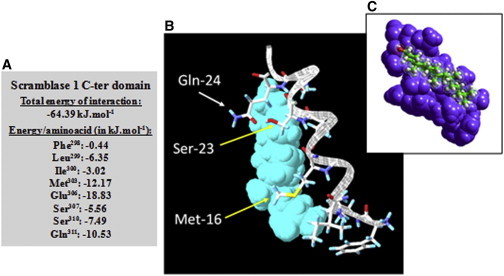
Human phospholipid scramblase 1 (SCR) catalyzes phospholipid transmembrane (flip-flop) motion. This protein is assumed to bind the membrane hydrophobic core through a transmembrane domain (TMD) as well as via covalently bound palmitoyl residues. Here, we explore the possible interaction of the SCR TMD with cholesterol by using a variety of experimental and computational biophysical approaches. Our findings indicate that SCR contains an amino acid segment at the C-terminal region that shows a remarkable affinity for cholesterol, although it lacks the CRAC sequence. Other 3-OH sterols, but not steroids lacking the 3-OH group, also bind this region of the protein. The newly identified cholesterol-binding region is located partly at the C-terminal portion of the TMD and partly in the first amino acid residues in the SCR C-terminal extracellular coil. This finding could be related to the previously described affinity of SCR for cholesterol-rich domains in membranes.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Bassé F., Stout J.G., Wiedmer T. Isolation of an erythrocyte membrane protein that mediates Ca2+-dependent transbilayer movement of phospholipid. J. Biol. Chem. 1996;271:17205–17210. - PubMed
-
- Zhou Q., Zhao J., Sims P.J. Molecular cloning of human plasma membrane phospholipid scramblase. A protein mediating transbilayer movement of plasma membrane phospholipids. J. Biol. Chem. 1997;272:18240–18244. - PubMed
-
- Sahu S.K., Gummadi S.N., Aradhyam G.K. Phospholipid scramblases: an overview. Arch. Biochem. Biophys. 2007;462:103–114. - PubMed
-
- Ben-Efraim I., Zhou Q., Sims P.J. Phospholipid scramblase 1 is imported into the nucleus by a receptor-mediated pathway and interacts with DNA. Biochemistry. 2004;43:3518–3526. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
