

In vivo orientation of single myosin lever arms in zebrafish skeletal muscle
- PMID: 25229148
- PMCID: PMC4167300
- DOI: 10.1016/j.bpj.2014.07.055
In vivo orientation of single myosin lever arms in zebrafish skeletal muscle
Abstract
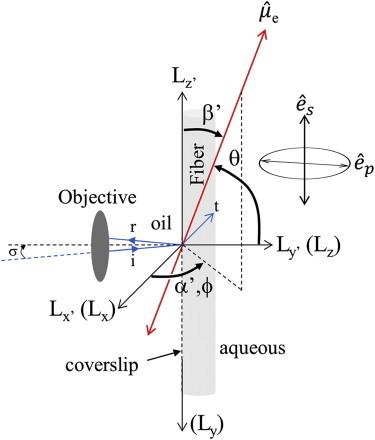
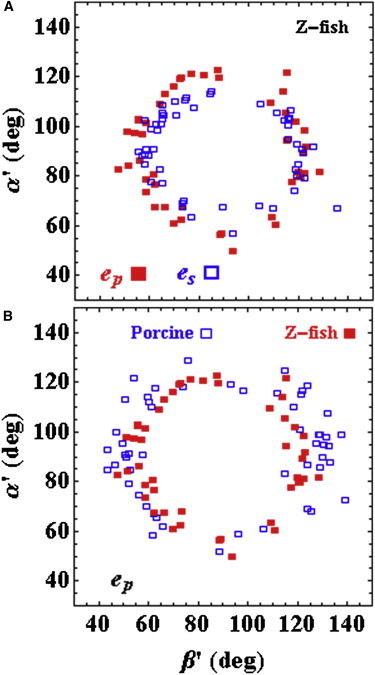
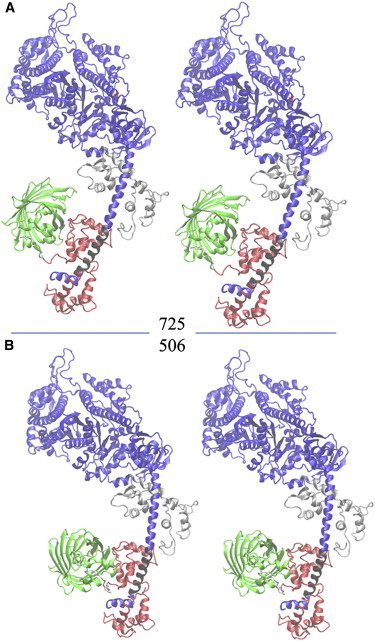
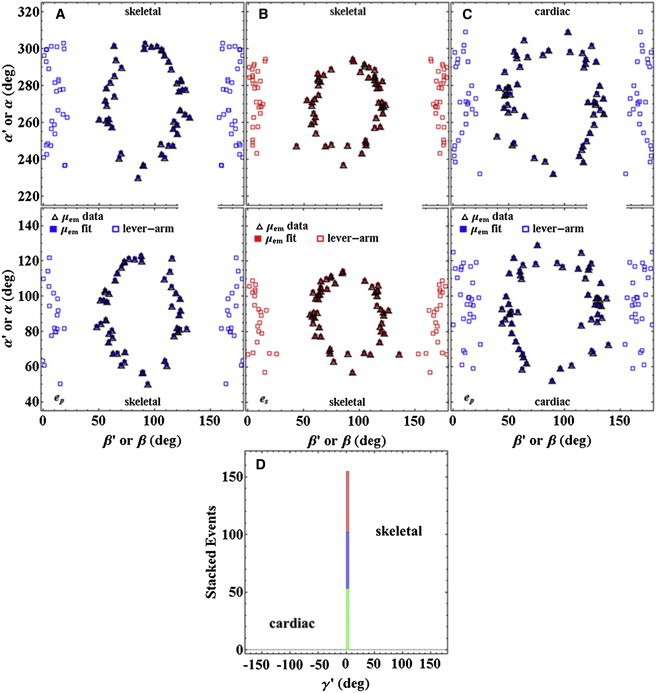
Cardiac and skeletal myosin assembled in the muscle lattice power contraction by transducing ATP free energy into the mechanical work of moving actin. Myosin catalytic/lever-arm domains comprise the transduction/mechanical coupling machinery that move actin by lever-arm rotation. In vivo, myosin is crowded and constrained by the fiber lattice as side chains are mutated and otherwise modified under normal, diseased, or aging conditions that collectively define the native myosin environment. Single-myosin detection uniquely defines bottom-up characterization of myosin functionality. The marriage of in vivo and single-myosin detection to study zebrafish embryo models of human muscle disease is a multiscaled technology that allows one-to-one registration of a selected myosin molecular alteration with muscle filament-sarcomere-cell-fiber-tissue-organ- and organism level phenotypes. In vivo single-myosin lever-arm orientation was observed at superresolution using a photoactivatable-green-fluorescent-protein (PAGFP)-tagged myosin light chain expressed in zebrafish skeletal muscle. By simultaneous observation of multiphoton excitation fluorescence emission and second harmonic generation from myosin, we demonstrated tag specificity for the lever arm. Single-molecule detection used highly inclined parallel beam illumination and was verified by quantized photoactivation and photobleaching. Single-molecule emission patterns from relaxed muscle in vivo provided extensive superresolved dipole orientation constraints that were modeled using docking scenarios generated for the myosin (S1) and GFP crystal structures. The dipole orientation data provided sufficient constraints to estimate S1/GFP coordination. The S1/GFP coordination in vivo is rigid and the lever-arm orientation distribution is well-ordered in relaxed muscle. For comparison, single myosins in relaxed permeabilized porcine papillary muscle fibers indicated slightly differently oriented lever arms and rigid S1/GFP coordination. Lever arms in both muscles indicated one preferred spherical polar orientation and widely distributed azimuthal orientations relative to the fiber symmetry axis. Cardiac myosin is more radially displaced from the fiber axis. Probe rigidity implies the PAGFP tag reliably indicates cross-bridge orientation in situ and in vivo.
Copyright © 2014 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures



References
-
- Lowey S., Waller G.S., Trybus K.M. Function of skeletal muscle myosin heavy and light chain isoforms by an in vitro motility assay. J. Biol. Chem. 1993;268:20414–20418. - PubMed
-
- Oldfors A. Hereditary myosin myopathies. Neuromuscul. Disord. 2007;17:355–367. - PubMed
-
- Toydemir R.M., Rutherford A., Bamshad M.J. Mutations in embryonic myosin heavy chain (MYH3) cause Freeman-Sheldon syndrome and Sheldon-Hall syndrome. Nat. Genet. 2006;38:561–565. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
