

Elephant endotheliotropic herpesviruses EEHV1A, EEHV1B, and EEHV2 from cases of hemorrhagic disease are highly diverged from other mammalian herpesviruses and may form a new subfamily
- PMID: 25231303
- PMCID: PMC4248956
- DOI: 10.1128/JVI.01673-14
Elephant endotheliotropic herpesviruses EEHV1A, EEHV1B, and EEHV2 from cases of hemorrhagic disease are highly diverged from other mammalian herpesviruses and may form a new subfamily
Abstract
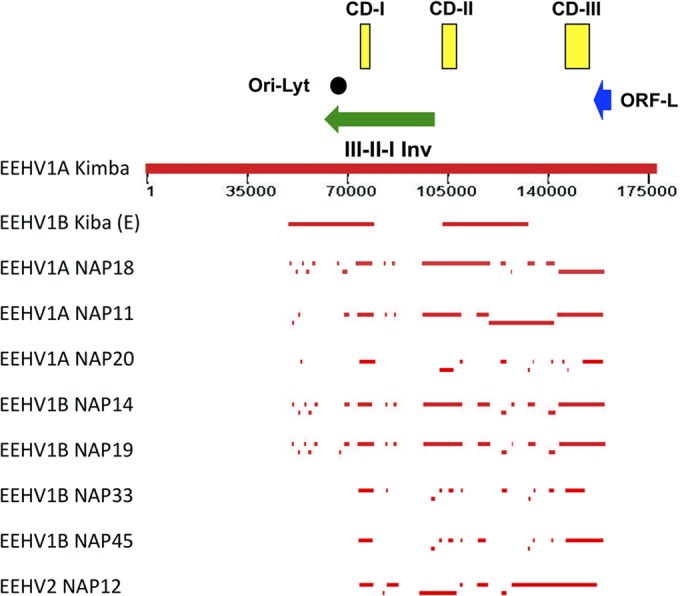
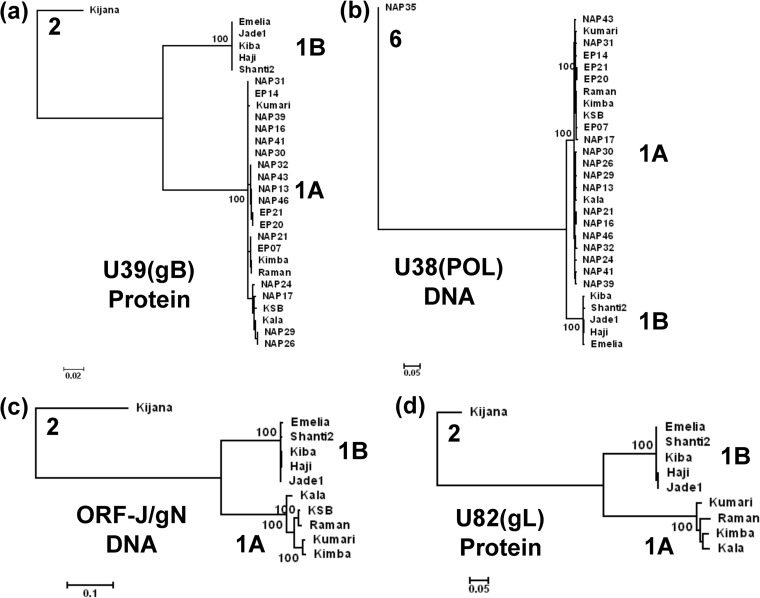
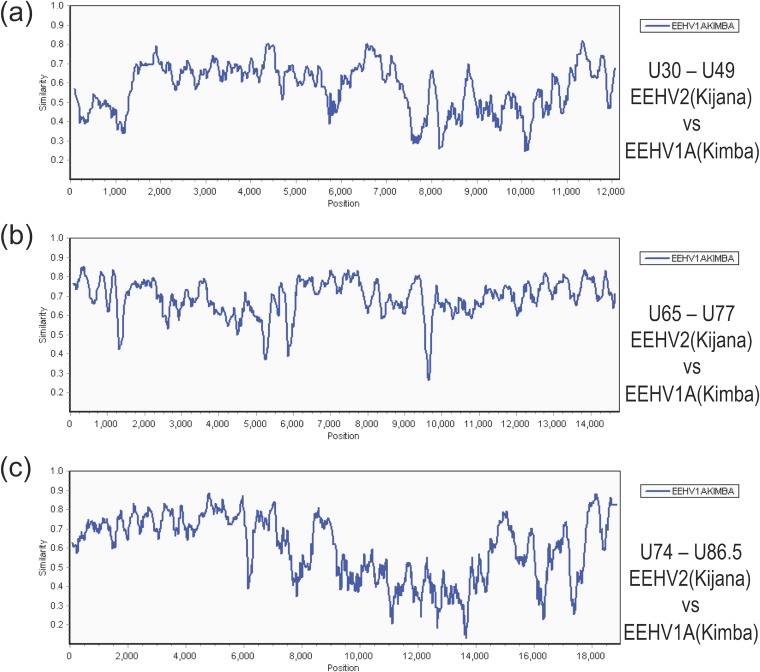
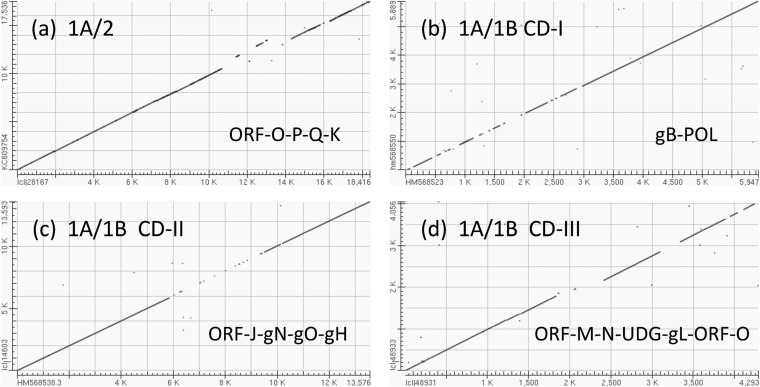
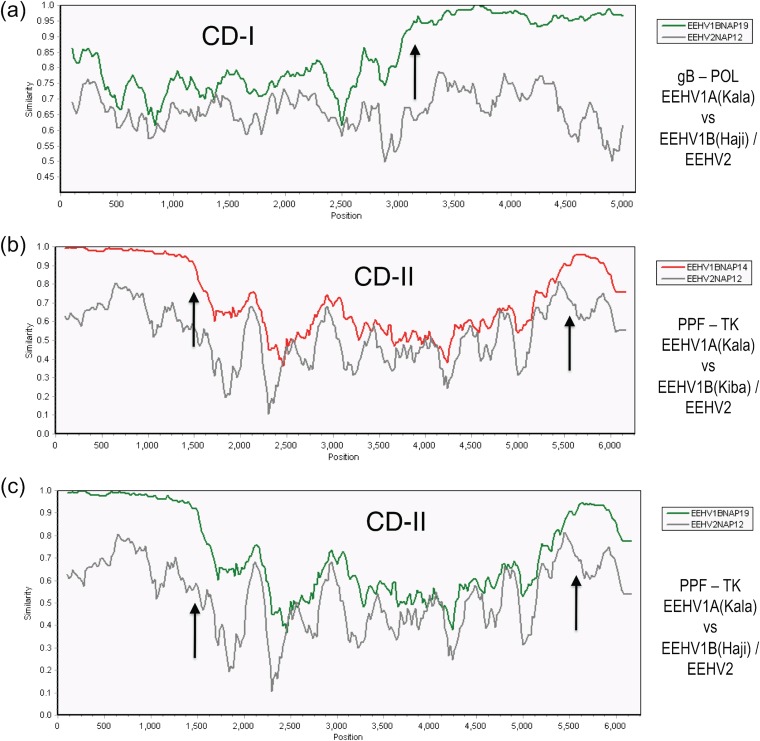
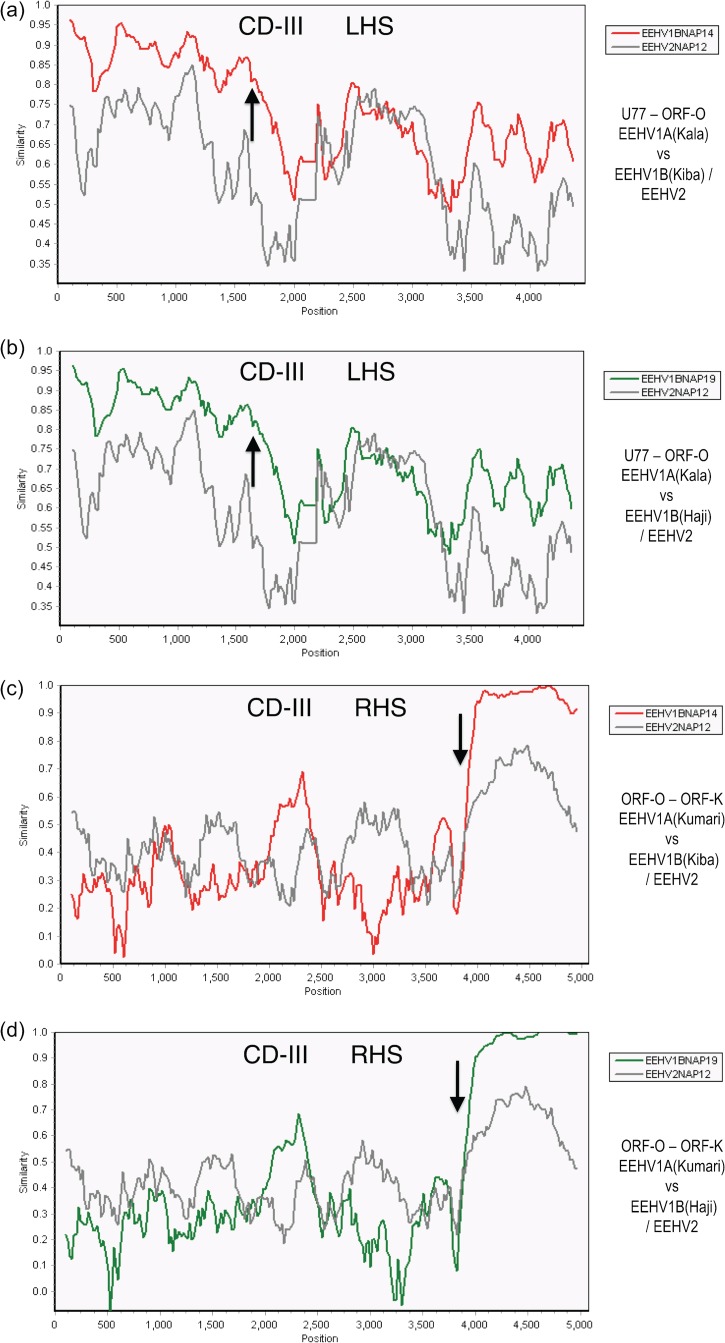
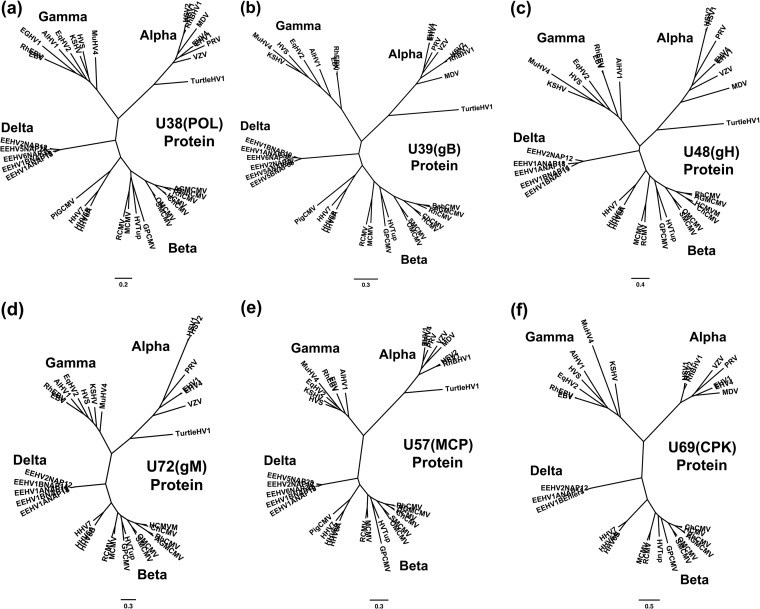
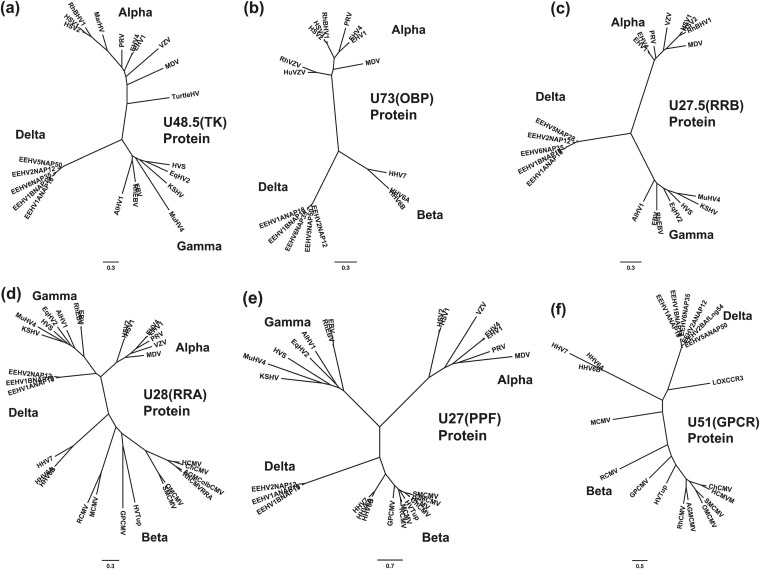
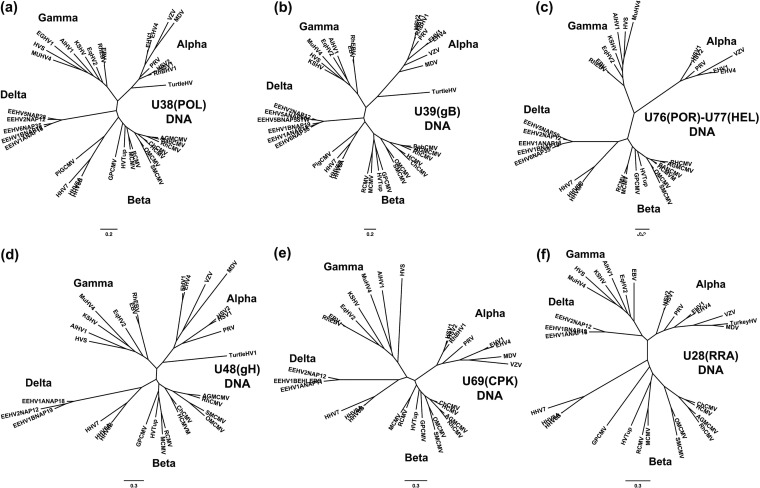
A family of novel endotheliotropic herpesviruses (EEHVs) assigned to the genus Proboscivirus have been identified as the cause of fatal hemorrhagic disease in 70 young Asian elephants worldwide. Although EEHV cannot be grown in cell culture, we have determined a total of 378 kb of viral genomic DNA sequence directly from clinical tissue samples from six lethal cases and two survivors. Overall, the data obtained encompass 57 genes, including orthologues of 32 core genes common to all herpesviruses, 14 genes found in some other herpesviruses, plus 10 novel genes, including a single large putative transcriptional regulatory protein (ORF-L). On the basis of differences in gene content and organization plus phylogenetic analyses of conserved core proteins that have just 20% to 50% or less identity to orthologues in other herpesviruses, we propose that EEHV1A, EEHV1B, and EEHV2 could be considered a new Deltaherpesvirinae subfamily of mammalian herpesviruses that evolved as an intermediate branch between the Betaherpesvirinae and Gammaherpesvirinae. Unlike cytomegaloviruses, EEHV genomes encode ribonucleotide kinase B subunit (RRB), thymidine kinase (TK), and UL9-like origin binding protein (OBP) proteins and have an alphaherpesvirus-like dyad symmetry Ori-Lyt domain. They also differ from all known betaherpesviruses by having a 40-kb large-scale inversion of core gene blocks I, II, and III. EEHV1 and EEHV2 DNA differ uniformly by more than 25%, but EEHV1 clusters into two major subgroups designated EEHV1A and EEHV1B with ancient partially chimeric features. Whereas large segments are nearly identical, three nonadjacent loci totaling 15 kb diverge by between 21 and 37%. One strain of EEHV1B analyzed is interpreted to be a modern partial recombinant with EEHV1A.
Importance: Asian elephants are an endangered species whose survival is under extreme pressure in wild range countries and whose captive breeding populations in zoos are not self-sustaining. In 1999, a novel class of herpesviruses called EEHVs was discovered. These viruses have caused a rapidly lethal hemorrhagic disease in 20% of all captive Asian elephant calves born in zoos in the United States and Europe since 1980. The disease is increasingly being recognized in Asian range countries as well. These viruses cannot be grown in cell culture, but by direct PCR DNA sequence analysis from segments totaling 15 to 30% of the genomes from blood or necropsy tissue from eight different cases, we have determined that they fall into multiple types and chimeric subtypes of a novel Proboscivirus genus, and we propose that they should also be classified as the first examples of a new mammalian herpesvirus subfamily named the Deltaherpesvirinae.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
Comment in
-
Trunkloads of viruses.J Virol. 2014 Dec;88(23):13520-2. doi: 10.1128/JVI.02359-14. Epub 2014 Sep 17. J Virol. 2014. PMID: 25231304 Free PMC article.
Similar articles
-
Review of Elephant Endotheliotropic Herpesviruses and Acute Hemorrhagic Disease.ILAR J. 2016;56(3):283-96. doi: 10.1093/ilar/ilv041. ILAR J. 2016. PMID: 26912715 Free PMC article. Review.
-
Comparative genome analysis of four elephant endotheliotropic herpesviruses, EEHV3, EEHV4, EEHV5, and EEHV6, from cases of hemorrhagic disease or viremia.J Virol. 2014 Dec;88(23):13547-69. doi: 10.1128/JVI.01675-14. Epub 2014 Sep 17. J Virol. 2014. PMID: 25231309 Free PMC article.
-
Detection of Quiescent Infections with Multiple Elephant Endotheliotropic Herpesviruses (EEHVs), Including EEHV2, EEHV3, EEHV6, and EEHV7, within Lymphoid Lung Nodules or Lung and Spleen Tissue Samples from Five Asymptomatic Adult African Elephants.J Virol. 2015 Dec 30;90(6):3028-43. doi: 10.1128/JVI.02936-15. J Virol. 2015. PMID: 26719245 Free PMC article.
-
Detection and evaluation of novel herpesviruses in routine and pathological samples from Asian and African elephants: identification of two new probosciviruses (EEHV5 and EEHV6) and two new gammaherpesviruses (EGHV3B and EGHV5).Vet Microbiol. 2011 Jan 10;147(1-2):28-41. doi: 10.1016/j.vetmic.2010.05.042. Epub 2010 Jun 1. Vet Microbiol. 2011. PMID: 20579821 Free PMC article.
-
The DNA sequence of human herpesvirus-6: structure, coding content, and genome evolution.Virology. 1995 May 10;209(1):29-51. doi: 10.1006/viro.1995.1228. Virology. 1995. PMID: 7747482 Review.
Cited by
-
Elephant Endotheliotropic Herpesvirus Hemorrhagic Disease in Asian Elephant Calves in Logging Camps, Myanmar.Emerg Infect Dis. 2020 Jan;26(1):63-69. doi: 10.3201/eid2601.190159. Emerg Infect Dis. 2020. PMID: 31855135 Free PMC article.
-
Pharmacokinetics and analytical determination of acyclovir in Asian elephant calves (Elephas maximus).Vet Anim Sci. 2021 Dec 24;15:100227. doi: 10.1016/j.vas.2021.100227. eCollection 2022 Mar. Vet Anim Sci. 2021. PMID: 35024493 Free PMC article.
-
Production of antibody against elephant endotheliotropic herpesvirus (EEHV) unveils tissue tropisms and routes of viral transmission in EEHV-infected Asian elephants.Sci Rep. 2018 Mar 16;8(1):4675. doi: 10.1038/s41598-018-22968-5. Sci Rep. 2018. PMID: 29549315 Free PMC article.
-
Identification of shedders of elephant endotheliotropic herpesviruses among Asian elephants (Elephas maximus) in Switzerland.PLoS One. 2017 May 3;12(5):e0176891. doi: 10.1371/journal.pone.0176891. eCollection 2017. PLoS One. 2017. PMID: 28467495 Free PMC article.
-
Review of Elephant Endotheliotropic Herpesviruses and Acute Hemorrhagic Disease.ILAR J. 2016;56(3):283-96. doi: 10.1093/ilar/ilv041. ILAR J. 2016. PMID: 26912715 Free PMC article. Review.
References
-
- Jacobson ER, Sundberg JP, Gaskin JM, Kollias GV, O'Banion MK. 1986. Cutaneous papillomas associated with a herpesvirus-like infection in a herd of captive African elephants. J. Am. Vet. Med. Assoc. 189:1075–1078. - PubMed
-
- McCully RM, Basson PA, Pienaar JG, Erasmus BJ, Young E. 1971. Herpes nodules in the lung of the African elephant (Loxodonta africana (Blumebach, 1792)). Onderstepoort J. Vet. Res. 38:225–235. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
